

The tyrosine phosphatase PTPRO sensitizes colon cancer cells to anti-EGFR therapy through activation of SRC-mediated EGFR signaling
- PMID: 25301722
- PMCID: PMC4259406
- DOI: 10.18632/oncotarget.2458
The tyrosine phosphatase PTPRO sensitizes colon cancer cells to anti-EGFR therapy through activation of SRC-mediated EGFR signaling
Abstract
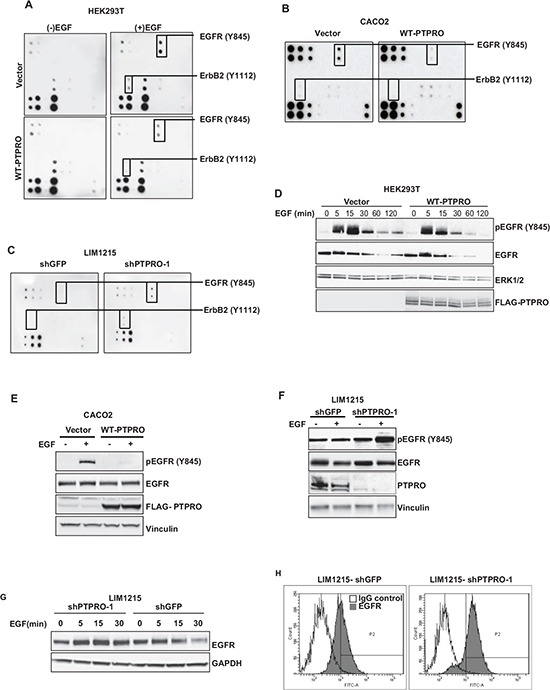
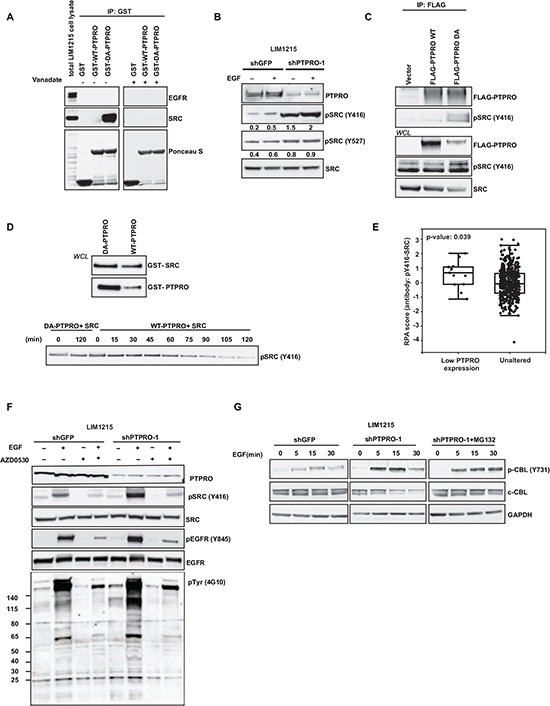
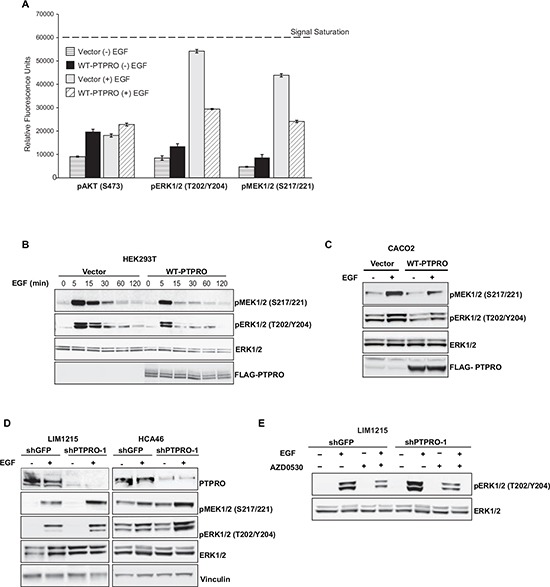
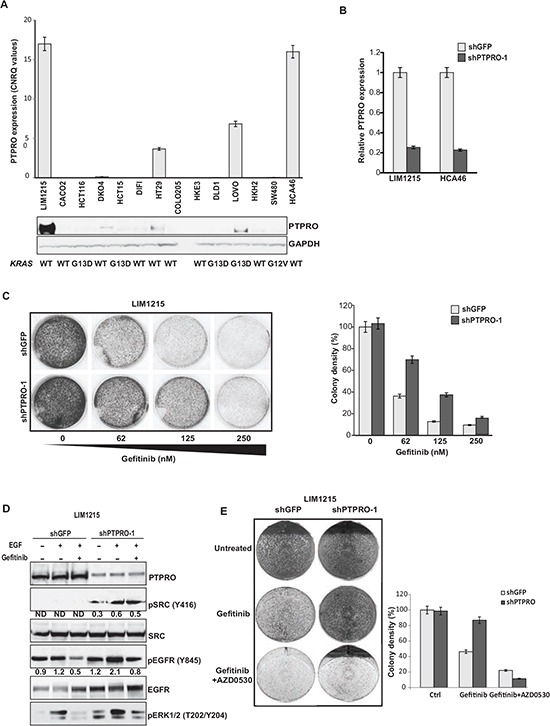
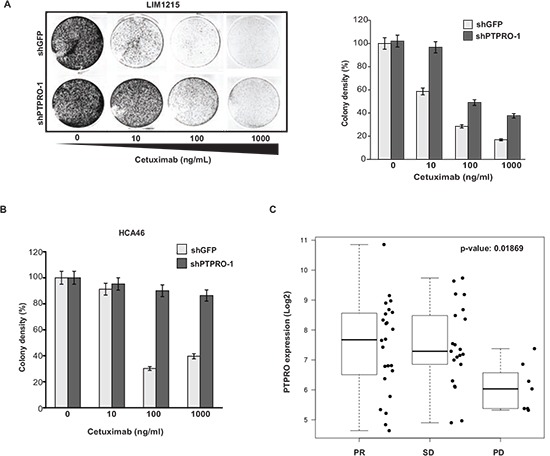
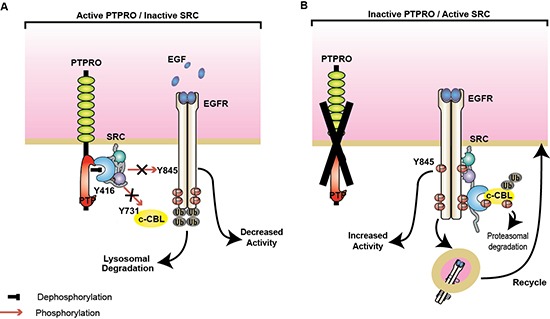
Inappropriate activation of epidermal growth factor receptor (EGFR) plays a causal role in many cancers including colon cancer. The activation of EGFR by phosphorylation is balanced by receptor kinase and protein tyrosine phosphatase activities. However, the mechanisms of negative EGFR regulation by tyrosine phosphatases remain largely unexplored. Our previous results indicate that protein tyrosine phosphatase receptor type O (PTPRO) is down-regulated in a subset of colorectal cancer (CRC) patients with a poor prognosis. Here we identified PTPRO as a phosphatase that negatively regulates SRC by directly dephosphorylating Y416 phosphorylation site. SRC activation triggered by PTPRO down-regulation induces phosphorylation of both EGFR at Y845 and the c-CBL ubiquitin ligase at Y731. Increased EGFR phosphorylation at Y845 promotes its receptor activity, whereas enhanced phosphorylation of c-CBL triggers its degradation promoting EGFR stability. Importantly, hyperactivation of SRC/EGFR signaling triggered by loss of PTPRO leads to high resistance of colon cancer to EGFR inhibitors. Our results not only highlight the PTPRO contribution in negative regulation of SRC/EGFR signaling but also suggest that tumors with low PTPRO expression may be therapeutically targetable by anti-SRC therapies.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Avraham R, Yarden Y. Feedback regulation of EGFR signalling: decision making by early and delayed loops. Nat Rev Mol Cell Biol. 2011;12:104–117. - PubMed
-
- Citri A, Yarden Y. EGF-ERBB signalling: towards the systems level. Nat Rev Mol Cell Biol. 2006;7:505–516. - PubMed
-
- Baselga J, Arteaga CL. Critical update and emerging trends in epidermal growth factor receptor targeting in cancer. J Clin Oncol. 2005;23:2445–2459. - PubMed
-
- Yarden Y, Pines G. The ERBB network: at last, cancer therapy meets systems biology. Nat Rev Cancer. 2012;12:553–563. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
