

Review
doi: 10.1016/j.cell.2014.09.016.
Cellular differences in protein synthesis regulate tissue homeostasis
Affiliations
- PMID: 25303523
- PMCID: PMC4222182
- DOI: 10.1016/j.cell.2014.09.016
Item in Clipboard
Review
Cellular differences in protein synthesis regulate tissue homeostasis
Cell.
.
Abstract
Although sometimes considered a "house-keeping" function, multiple aspects of protein synthesis are regulated differently among somatic cells, including stem cells, and can be modulated in a cell-type-specific manner. These differences are required to establish and maintain differences in cell identity, cell function, tissue homeostasis, and tumor suppression.
Copyright © 2014 Elsevier Inc. All rights reserved.
Figures
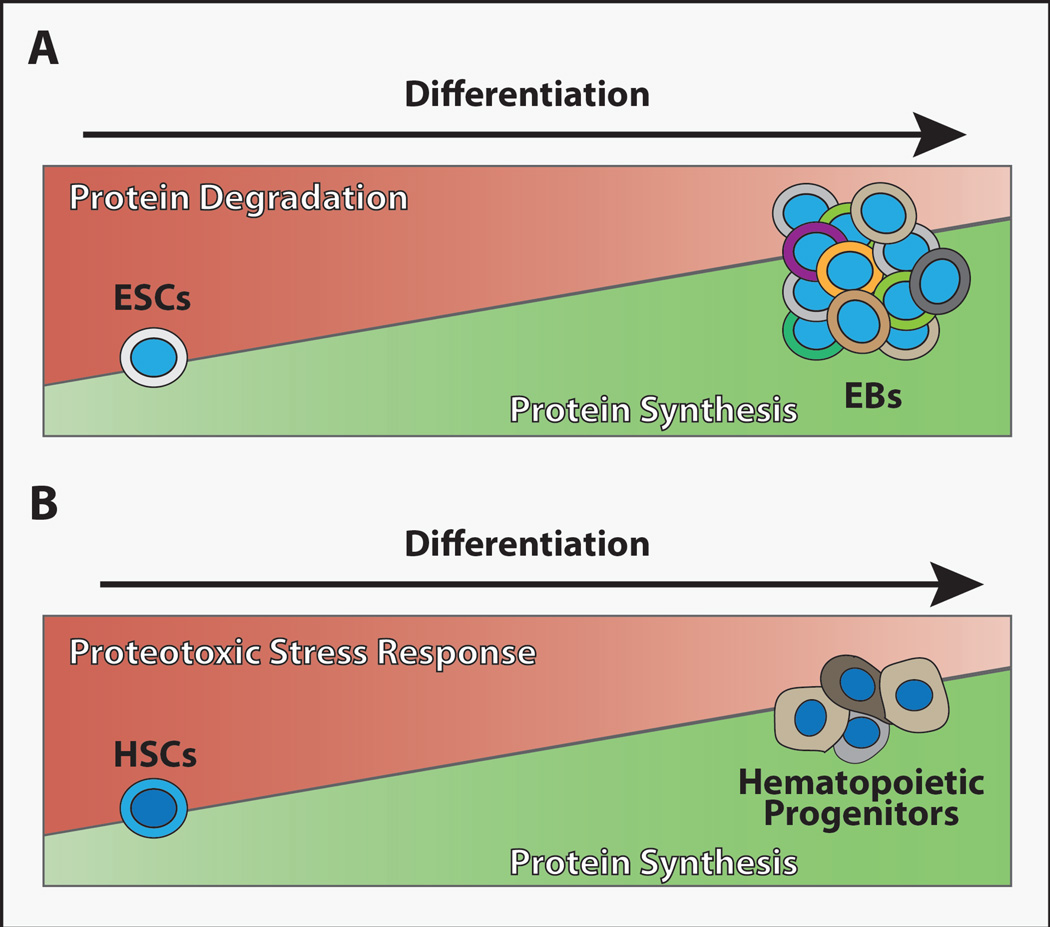
(A) Global protein translation levels remain low in embryonic stem cells (ESCs) but increase as these cells differentiate to form embryoid bodies (EBs) (Ingolia et al., 2011; Sampath et al., 2008). By contrast, the activity of protein degradation mechanisms appears high in ESCs relative to EBs (Vilchez et al., 2012). (B) Adult HSCs display low levels of global protein synthesis relative to differentiating hematopoietic progenitor cells (Signer et al., 2014), and proteotoxic stress response appears enhanced in umbilical cord blood (fetal) HSCs (van Galen et al., 2014). Whether these trends in proteostasis regulatory mechanisms will be seen in other stem cell populations remains unclear, but keeping overall protein levels low in long-lived stem cell populations may promote their fitness and longevity.
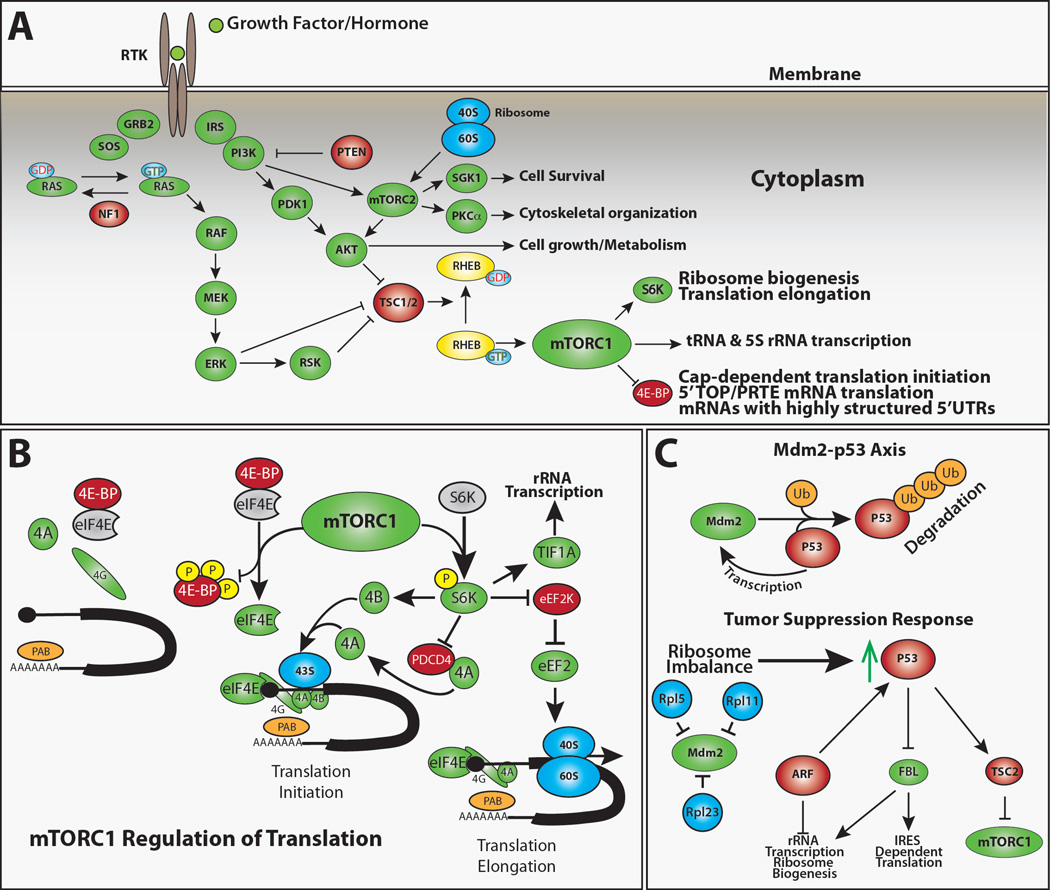
(A) Growth factors, hormones, nutrients, and stress all influence protein synthesis through complex signaling pathways (Laplante and Sabatini, 2012; Topisirovic and Sonenberg, 2011). mTORC1 preferentially promotes the translation of a subset of mRNAs that contain complex 5’UTRs and clusters of 5’ terminal oligopyrimidines (TOP) or similar pyrimidine-rich sequences (Hsieh et al., 2012; Jefferies et al., 1994; Thoreen et al., 2012). These TOP genes primarily encode proteins that promote translation, including ribosomal proteins, initiation factors, and elongation factors (Thoreen et al., 2012). While many proteins have been proposed to regulate the translation of TOP mRNAs (Damgaard and Lykke-Andersen, 2011; Tcherkezian et al., 2014) and the translation of specific TOP containing mRNAs is regulated in a cell-specific manner (Avni et al., 1997; Ivanov et al., 2011), the roles of these elements in stem cell maintenance and tissue homeostasis remain unclear. (B) The regulation of translation initiation represents the major mechanism by which cells regulate protein synthesis. The eIF4F complex, which includes eIF4E, eIF4G and eIF4A, promotes the translational initiation of virtually all cellular mRNAs (Sonenberg and Hinnebusch, 2009) and mRNAs with highly structured 5’UTRs are particularly sensitive to eIF4E activity (Feoktistova et al., 2013; Koromilas et al., 1992). Activated mTORC1 phosphorylates 4E-BPs, preventing their inhibition of eIF4E. In the absence of 4E-BPs, eIF4E binds to the m7G mRNA cap and recruits eIF4G, which in turn acts as a scaffold for the RNA helicase eIF4A. The formation of this eIF4F complex on the 5’-end of mRNA promotes the initiation of cap-dependent translation. Activated mTORC1 also phosphorylates and activates S6K, which phosphorylates eIF4B, eEF2K and TIF1A. Phosphorylated eIF4B enhances the helicase activity of eIF4A to unwind secondary structures in mRNA, which promotes translational initiation. Phosphorylation of eEF2K by S6K interferes with its ability to block the activity of the translational elongation factor eEF2. Activation of TIF1A promotes ribosomal RNA transcription. (C) The E3 ubiquitin ligase Mdm2 promotes the degradation of the tumor suppressor p53. p53 acts in a feedback loop to promote the transcription of the Mdm2 gene. Thus under normal conditions, p53 levels remain low. Imbalances in ribosome biogenesis induce a tumor suppressor response in which free ribosomal proteins, particularly Rpl5 and Rpl11, bind and inhibit Mdm2 (Bhat et al., 2004; Dai and Lu, 2004; Dai et al., 2006; Zhang et al., 2003). This results in the accumulation of p53, which promotes the activity of TSC2, thereby repressing the activity of mTORC1. p53 and other tumor suppressors, such as ARF, inhibit rRNA transcription and ribosome biogenesis (Lessard et al., 2010; Sloan et al., 2013; Sugimoto et al., 2003).
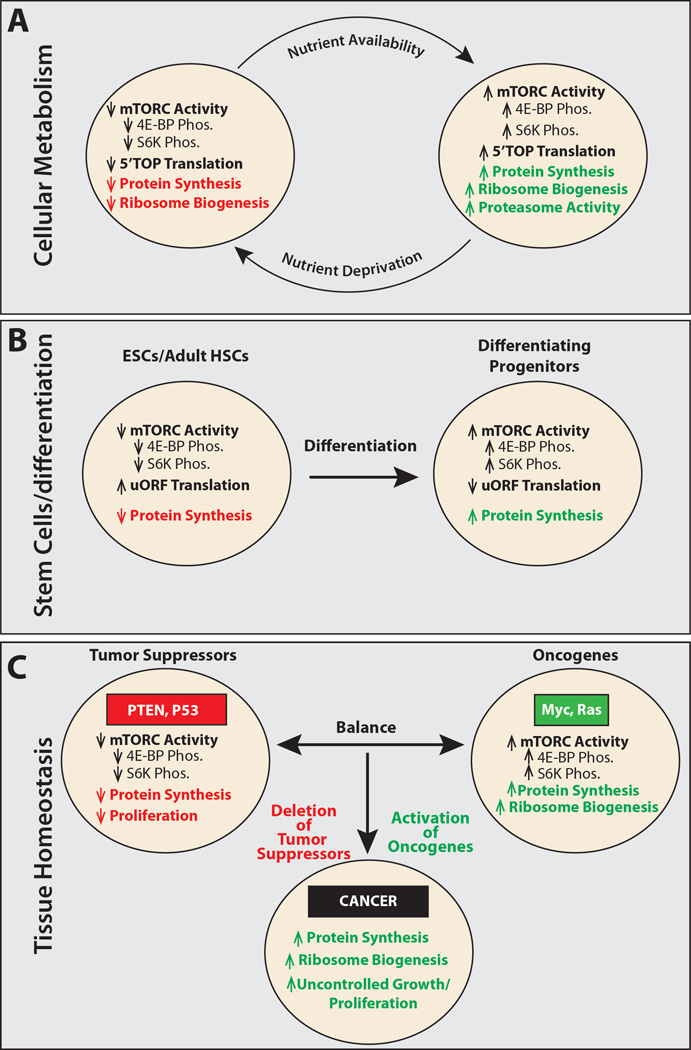
(A) Cells respond to changes in nutrient levels by altering protein synthesis, ribosome biogenesis (Laplante and Sabatini, 2012), and protein degradation (Zhang et al., 2014b) by modulating mTORC1 signaling. (B) Embryonic stem cells (ESCs) and adult hematopoietic stem cells (HSCs) exhibit lower levels of protein synthesis than their differentiating progeny. Whether other stem cell lineages display similar changes in protein synthesis upon differentiation will require further analysis. One intriguing possibility is that this property may vary among stem cells, depending on their cell cycle kinetics and whether they are long-lived or short-lived in vivo. (C) Tissue homeostasis depends on a balance between proto-oncogenes and tumor suppressors. Tumor suppressors such as PTEN and p53 reduce cellular growth and proliferation through a number of different mechanisms, including decreasing mTORC1 activity and reducing global protein synthesis (Laplante and Sabatini, 2012). Proto-oncogenes, including Myc and Ras, increase protein translation and ribosome biogenesis, fueling growth and proliferation. Cells transform to cancer when mutations reduce tumor suppressor function or increase oncogene function (van Riggelen et al., 2010).
References
-
- Abkowitz JL, Sabo KM, Nakamoto B, Blau CA, Martin FH, Zsebo KM, Papayannopoulou T. Diamond-blackfan anemia: in vitro response of erythroid progenitors to the ligand for c-kit. Blood. 1991;78:2198–2202. - PubMed
-
- Armistead J, Triggs-Raine B. Diverse diseases from a ubiquitous process: The ribosomopathy paradox. FEBS Letters. 2014;588:1491–1500. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
