

Increased resistance to biotrophic pathogens in the Arabidopsis constitutive induced resistance 1 mutant is EDS1 and PAD4-dependent and modulated by environmental temperature
- PMID: 25303634
- PMCID: PMC4193848
- DOI: 10.1371/journal.pone.0109853
Increased resistance to biotrophic pathogens in the Arabidopsis constitutive induced resistance 1 mutant is EDS1 and PAD4-dependent and modulated by environmental temperature
Abstract
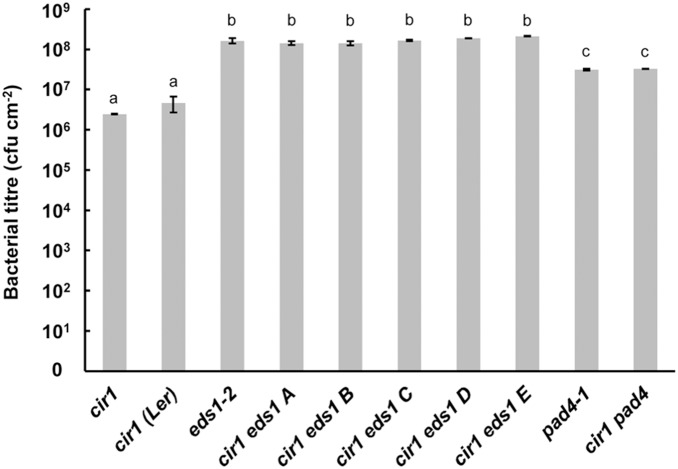
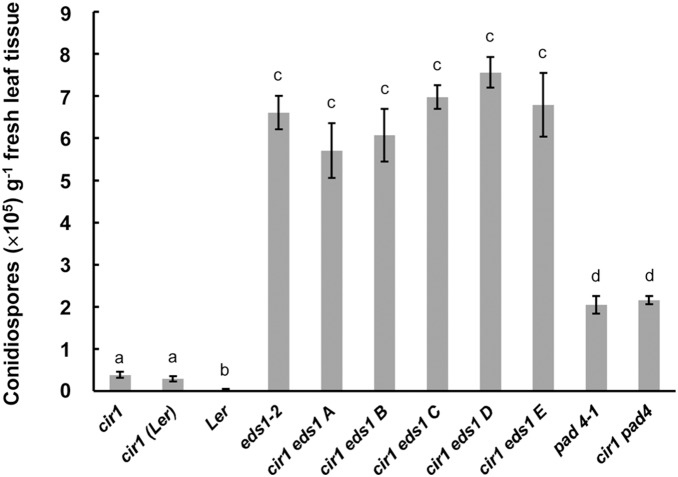
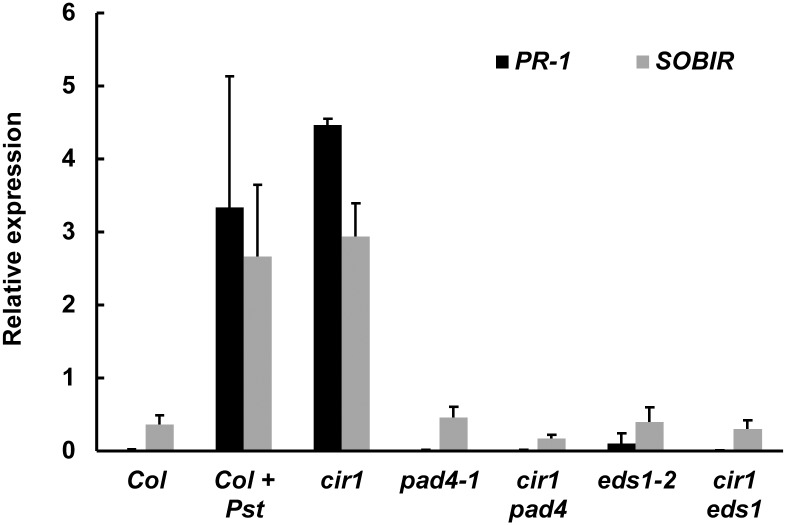
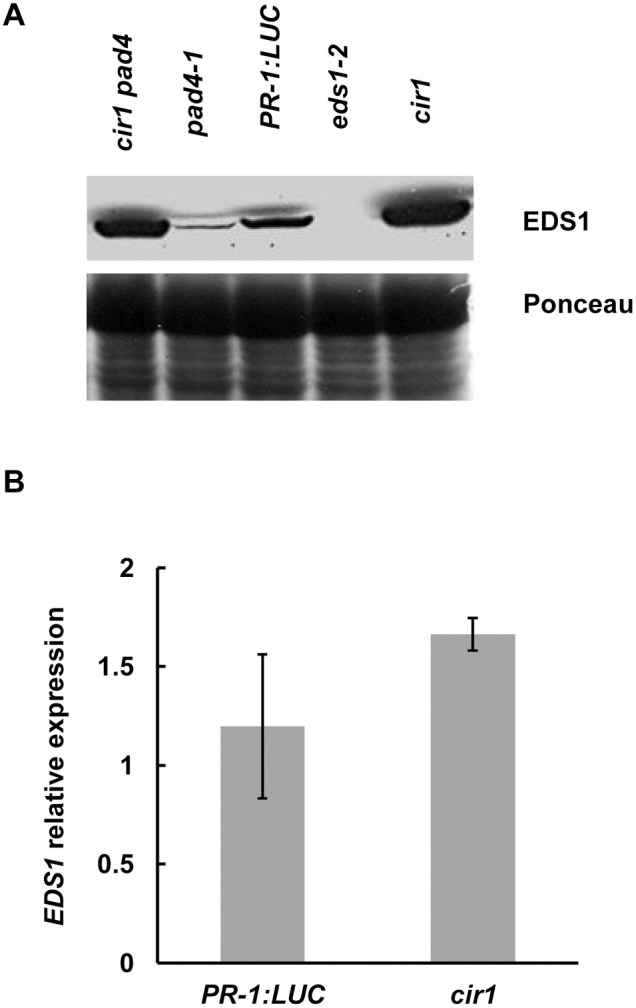
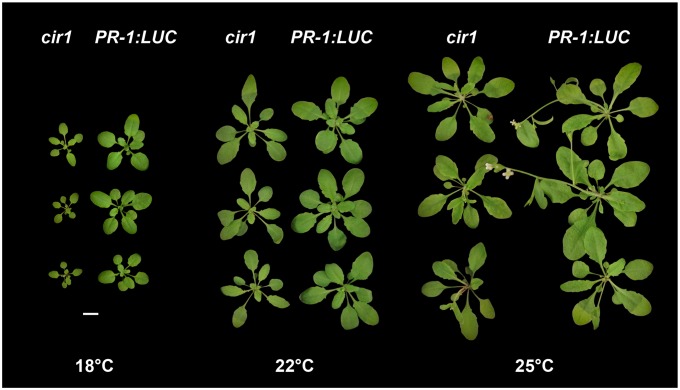
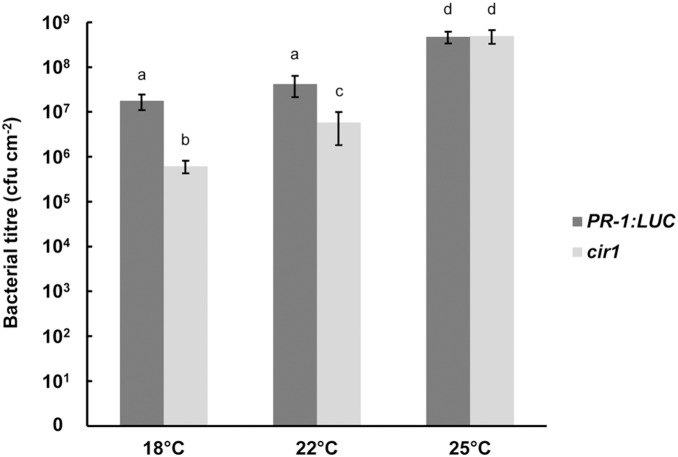
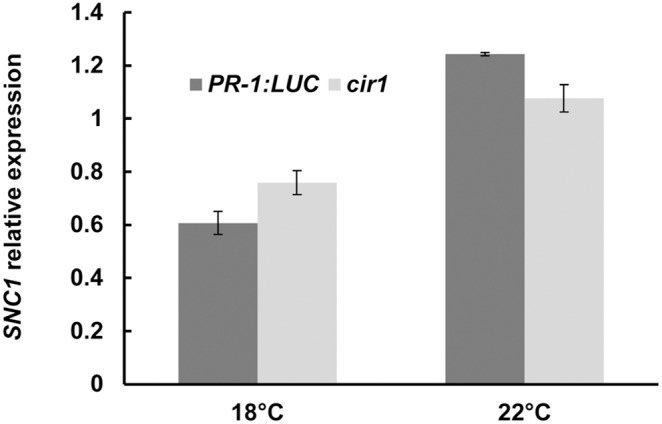
The Arabidopsis constitutive induced resistance 1 (cir1) mutant displays salicylic acid (SA)-dependent constitutive expression of defence genes and enhanced resistance to biotrophic pathogens. To further characterise the role of CIR1 in plant immunity we conducted epistasis analyses with two key components of the SA-signalling branch of the defence network, ENHANCED DISEASE SUSCEPTIBILITY1 (EDS1) and PHYTOALEXIN DEFICIENT4 (PAD4). We demonstrate that the constitutive defence phenotypes of cir1 require both EDS1 and PAD4, indicating that CIR1 lies upstream of the EDS1-PAD4 regulatory node in the immune signalling network. In light of this finding we examined EDS1 expression in cir1 and observed increased protein, but not mRNA levels in this mutant, suggesting that CIR1 might act as a negative regulator of EDS1 via a post-transcriptional mechanism. Finally, as environmental temperature is known to influence the outcome of plant-pathogen interactions, we analysed cir1 plants grown at 18, 22 or 25°C. We found that susceptibility to Pseudomonas syringae pv. tomato (Pst) DC3000 is modulated by temperature in cir1. Greatest resistance to this pathogen (relative to PR-1:LUC control plants) was observed at 18°C, while at 25°C no difference in susceptibility between cir1 and control plants was apparent. The increase in resistance to Pst DC3000 at 18°C correlated with a stunted growth phenotype, suggesting that activation of defence responses may be enhanced at lower temperatures in the cir1 mutant.
Conflict of interest statement
Figures
References
-
- Jones JD, Dangl JL (2006) The plant immune system. Nature 444: 323–329. - PubMed
-
- Schwessinger B, Ronald PC (2012) Plant innate immunity: perception of conserved microbial signatures. Ann Rev Plant Biol 63: 451–482. - PubMed
-
- Schwessinger B, Zipfel C (2008) News from the frontline: recent insights into PAMP-triggered immunity in plants. Curr Opin Plant Biol 11: 389–395. - PubMed
-
- Spoel SH, Dong X (2012) How do plants achieve immunity? Defence without specialized immune cells. Nature Rev Immunol 12: 89–100. - PubMed
-
- Fu ZQ, Dong X (2013) Systemic acquired resistance: turning local infection into global defense. Ann Rev Plant Biol 64: 839–863. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
