

HIV-1 Vif inhibits G to A hypermutations catalyzed by virus-encapsidated APOBEC3G to maintain HIV-1 infectivity
- PMID: 25304135
- PMCID: PMC4200127
- DOI: 10.1186/s12977-014-0089-5
HIV-1 Vif inhibits G to A hypermutations catalyzed by virus-encapsidated APOBEC3G to maintain HIV-1 infectivity
Abstract
Background: HIV-1 viral infectivity factor (Vif) is an essential accessory protein for HIV-1 replication. The predominant function of Vif is to counteract Apolipoprotein B mRNA-editing enzyme-catalytic polypeptide-like 3G (APOBEC3G, A3G), a potent host restriction factor that inhibits HIV-1 replication. Vif mediates the proteasomal degradation of A3G and inhibits A3G translation, thus diminishing the pool of A3G that is available to be packaged into budding virion. Although Vif is robust in degrading A3G, the protection provided against A3G is not absolute. Clinical and laboratory evidence have shown that A3G is not completely excluded from HIV-1 viral particles during HIV-1 replication. It remains unclear why the viral samples are still infectious when A3G has been packaged into the virions.
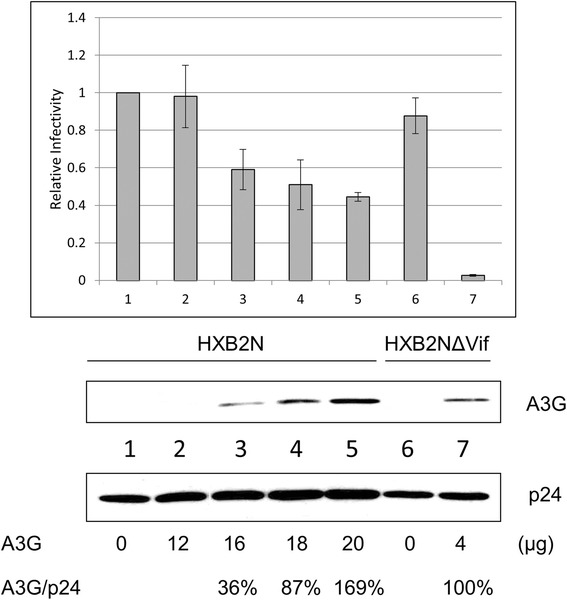
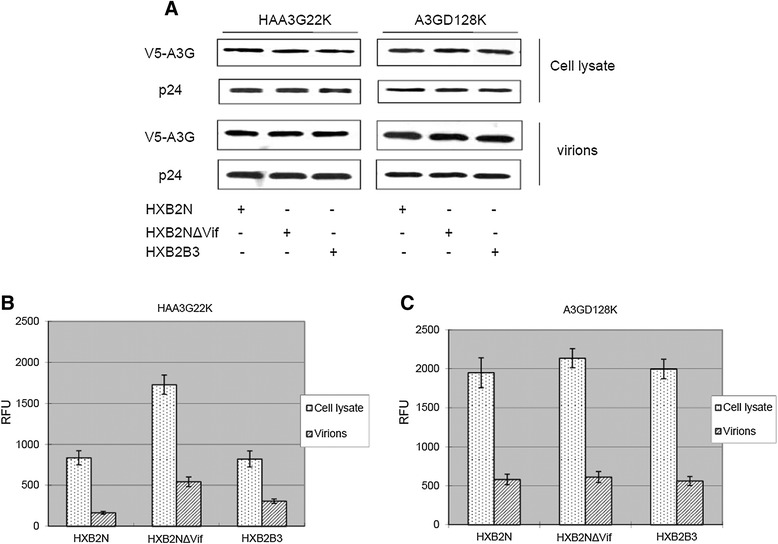
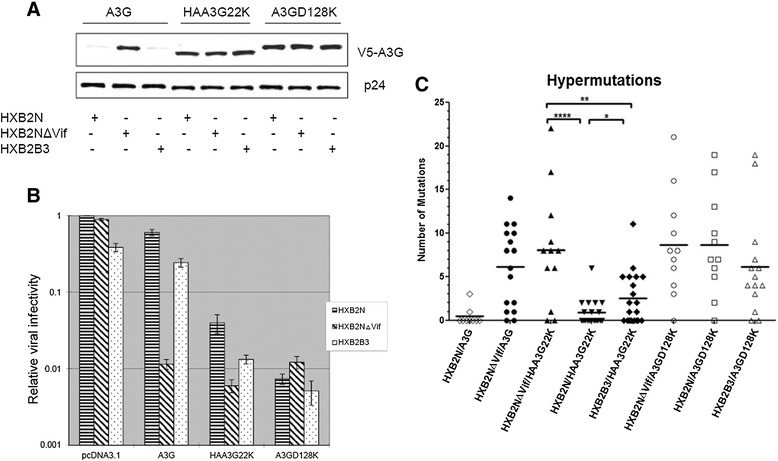
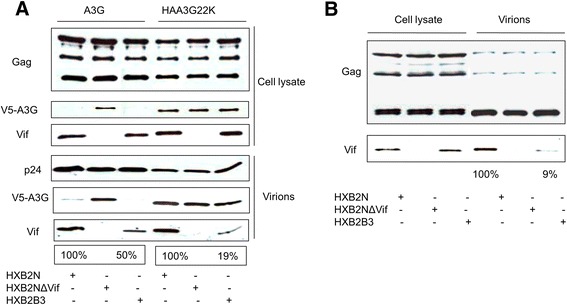
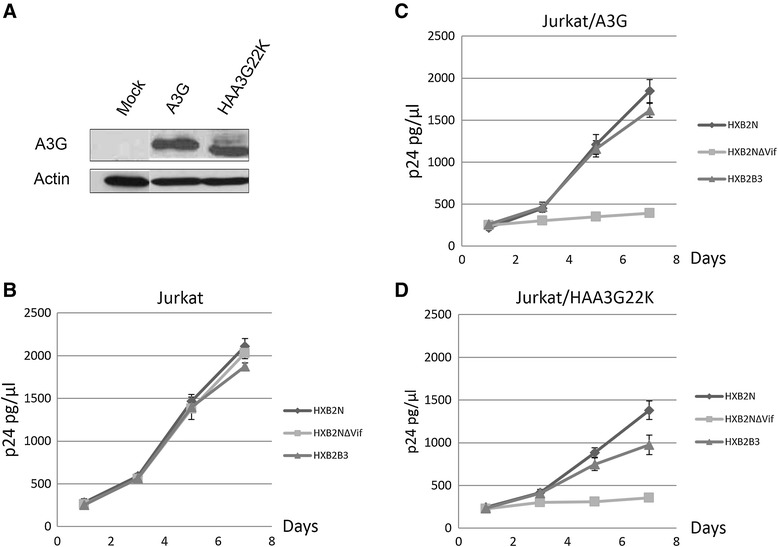
Results: In this study, we provide evidence that Vif continues to protect HIV-1 from the deleterious effects of A3G, even after packaging of A3G has occurred. When equal amounts of A3G were packaged into budding virions, the virus expressing functional Vif was more infectious and incurred fewer G to A hypermutations in the second round of infection compared to Vif-deficient virus. A Vif mutant with a defect in viral packaging showed a reduced ability to protect the HIV-1 genome from G to A hypermutations.
Conclusion: Our data suggest that even packaged A3G is still under the tyranny of Vif. Our work brings to light an additional caveat for any therapy that hopes to exploit the Vif-A3G axis. The ideal strategy would not only enhance A3G viral packaging, but also reduce HIV-1 Vif viral encapsidation.
Figures
Similar articles
-
Multifaceted counter-APOBEC3G mechanisms employed by HIV-1 Vif.J Mol Biol. 2011 Jul 29;410(5):1065-76. doi: 10.1016/j.jmb.2011.03.058. J Mol Biol. 2011. PMID: 21763507 Free PMC article.
-
GANP interacts with APOBEC3G and facilitates its encapsidation into the virions to reduce HIV-1 infectivity.J Immunol. 2013 Dec 15;191(12):6030-6039. doi: 10.4049/jimmunol.1302057. Epub 2013 Nov 6. J Immunol. 2013. PMID: 24198285 Free PMC article.
-
Mechanism of Enhanced HIV Restriction by Virion Coencapsidated Cytidine Deaminases APOBEC3F and APOBEC3G.J Virol. 2017 Jan 18;91(3):e02230-16. doi: 10.1128/JVI.02230-16. Print 2017 Feb 1. J Virol. 2017. PMID: 27881650 Free PMC article.
-
Various strategies for developing APOBEC3G protectors to circumvent human immunodeficiency virus type 1.Eur J Med Chem. 2023 Mar 15;250:115188. doi: 10.1016/j.ejmech.2023.115188. Epub 2023 Feb 6. Eur J Med Chem. 2023. PMID: 36773550 Review.
-
New insights into the role of Vif in HIV-1 replication.AIDS Rev. 2004 Jan-Mar;6(1):34-9. AIDS Rev. 2004. PMID: 15168739 Review.
Cited by
-
Dodging the Host Interferon-Stimulated Gene Mediated Innate Immunity by HIV-1: A Brief Update on Intrinsic Mechanisms and Counter-Mechanisms.Front Immunol. 2021 Jul 29;12:716927. doi: 10.3389/fimmu.2021.716927. eCollection 2021. Front Immunol. 2021. PMID: 34394123 Free PMC article. Review.
-
The Disassociation of A3G-Related HIV-1 cDNA G-to-A Hypermutation to Viral Infectivity.Viruses. 2024 May 4;16(5):728. doi: 10.3390/v16050728. Viruses. 2024. PMID: 38793610 Free PMC article.
-
Direct inhibition of human APOBEC3 deaminases by HIV-1 Vif independent of the proteolysis pathway.Biophys J. 2024 Feb 6;123(3):294-306. doi: 10.1016/j.bpj.2023.12.015. Epub 2023 Dec 19. Biophys J. 2024. PMID: 38115583 Free PMC article.
-
Degradation-Independent Inhibition of APOBEC3G by the HIV-1 Vif Protein.Viruses. 2021 Apr 3;13(4):617. doi: 10.3390/v13040617. Viruses. 2021. PMID: 33916704 Free PMC article. Review.
-
Tree Shrew Cells Transduced with Human CD4 and CCR5 Support Early Steps of HIV-1 Replication, but Viral Infectivity Is Restricted by APOBEC3.J Virol. 2021 Jul 26;95(16):e0002021. doi: 10.1128/JVI.00020-21. Epub 2021 Jul 26. J Virol. 2021. PMID: 34076481 Free PMC article.
References
-
- Lecossier D, Bouchonnet F, Clavel F, Hance AJ: Hypermutation of HIV-1 DNA in the absence of the Vif protein.Science 2003, 300:1112. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- T32 HL007737/HL/NHLBI NIH HHS/United States
- UL1 TR000445/TR/NCATS NIH HHS/United States
- UL1TR000445/TR/NCATS NIH HHS/United States
- G12 MD007586/MD/NIMHD NIH HHS/United States
- SC1 GM089269/GM/NIGMS NIH HHS/United States
- T32 AI007281/AI/NIAID NIH HHS/United States
- U54MD007593/MD/NIMHD NIH HHS/United States
- U54 MD007593/MD/NIMHD NIH HHS/United States
- R25 GM059994/GM/NIGMS NIH HHS/United States
- SC1GM089269/GM/NIGMS NIH HHS/United States
- P30AI054999/AI/NIAID NIH HHS/United States
- G12MD007586/MD/NIMHD NIH HHS/United States
- P30 AI054999/AI/NIAID NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
