

The neurotensin-HIF-1α-VEGFα axis orchestrates hypoxia, colonic inflammation, and intestinal angiogenesis
- PMID: 25307345
- PMCID: PMC4258496
- DOI: 10.1016/j.ajpath.2014.08.015
The neurotensin-HIF-1α-VEGFα axis orchestrates hypoxia, colonic inflammation, and intestinal angiogenesis
Abstract
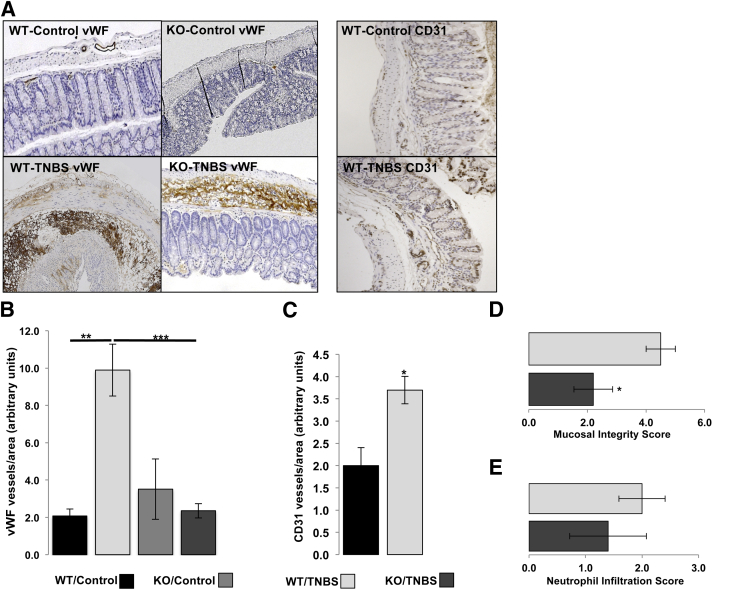
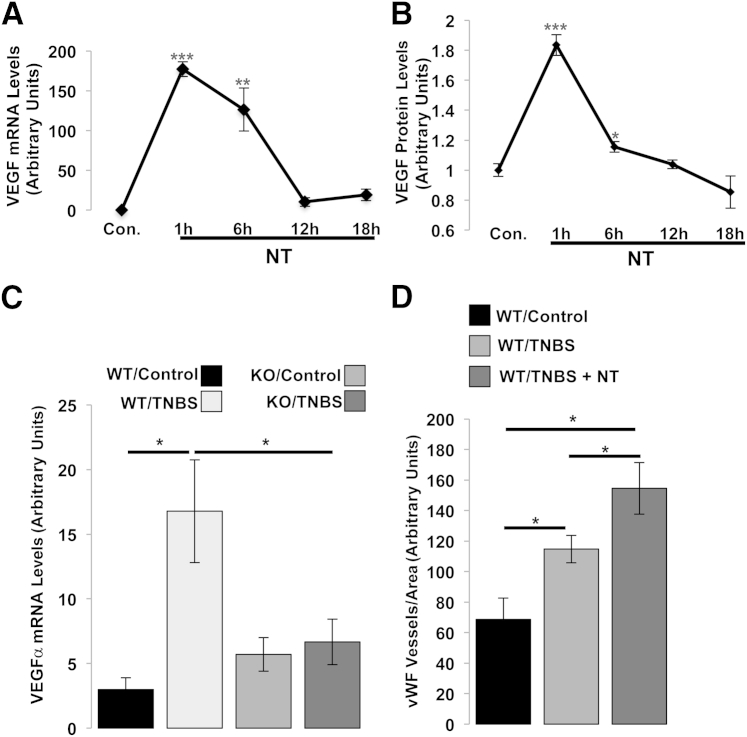
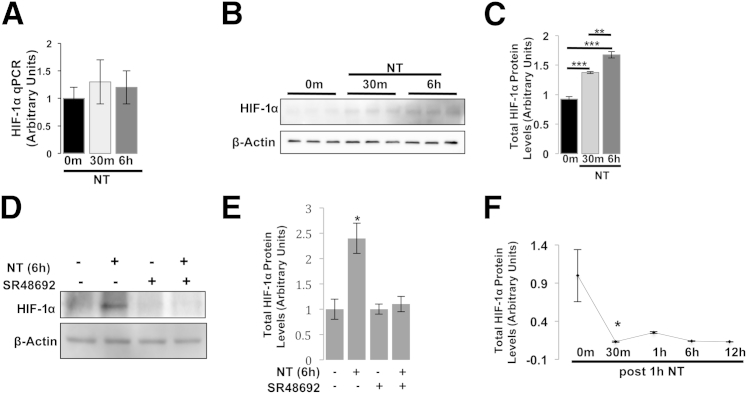
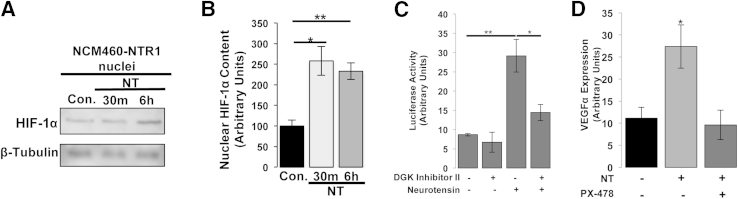
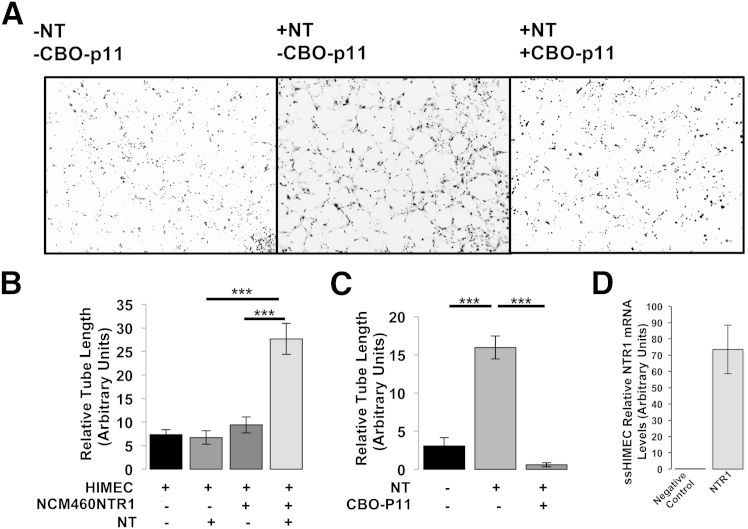
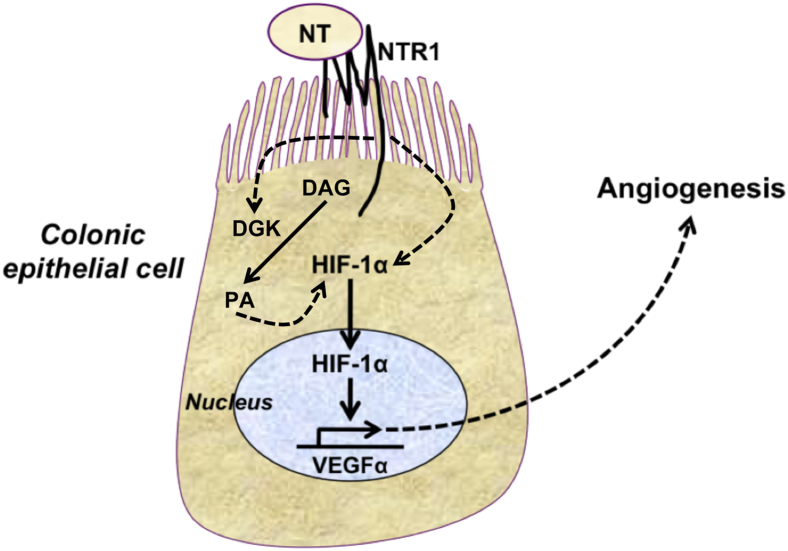
The expression of neurotensin (NT) and its receptor (NTR1) is up-regulated in experimental colitis and inflammatory bowel disease; NT/NTR1 interactions regulate gut inflammation. During active inflammation, metabolic shifts toward hypoxia lead to the activation of hypoxia-inducible factor (HIF)-1, which enhances vascular endothelial growth factor (VEGF) expression, promoting angiogenesis. We hypothesized that NT/NTR1 signaling regulates intestinal manifestations of hypoxia and angiogenesis by promoting HIF-1 transcriptional activity and VEGFα expression in experimental colitis. We studied NTR1 signaling in colitis-associated angiogenesis using 2,4,6-trinitrobenzenesulfonic acid-treated wild-type and NTR1-knockout mice. The effects of NT on HIF-1α and VEGFα were assessed on human colonic epithelial cells overexpressing NTR1 (NCM460-NTR1) and human intestinal microvascular-endothelial cells. NTR1-knockout mice had reduced microvascular density and mucosal integrity score compared with wild-type mice after 2,4,6-trinitrobenzenesulfonic acid treatment. VEGFα mRNA levels were increased in NCM460-NTR1 cells treated with 10(-7) mol/L NT, at 1 and 6 hours post-treatment. NT exposure in NCM460-NTR1 cells caused stabilization, nuclear translocation, and transcriptional activity of HIF-1α in a diacylglycerol kinase-dependent manner. NT did not stimulate tube formation in isolated human intestinal macrovascular endothelial cells but did so in human intestinal macrovascular endothelial cells cocultured with NCM460-NTR1 cells. Our results demonstrate the importance of an NTR1-HIF-1α-VEGFα axis in intestinal angiogenic responses and in the pathophysiology of colitis and inflammatory bowel disease.
Copyright © 2014 American Society for Investigative Pathology. Published by Elsevier Inc. All rights reserved.
Figures
References
-
- Carraway R., Leeman S.E. The isolation of a new hypotensive peptide, neurotensin, from bovine hypothalami. J Biol Chem. 1973;248:6854–6861. - PubMed
-
- Uhl G.R., Goodman R.R., Snyder S.H. Neurotensin-containing cell bodies, fibers and nerve terminals in the brain stem of the rat: immunohistochemical mapping. Brain Res. 1979;167:77–91. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
