

The beginning of a seed: regulatory mechanisms of double fertilization
- PMID: 25309552
- PMCID: PMC4160995
- DOI: 10.3389/fpls.2014.00452
The beginning of a seed: regulatory mechanisms of double fertilization
Abstract
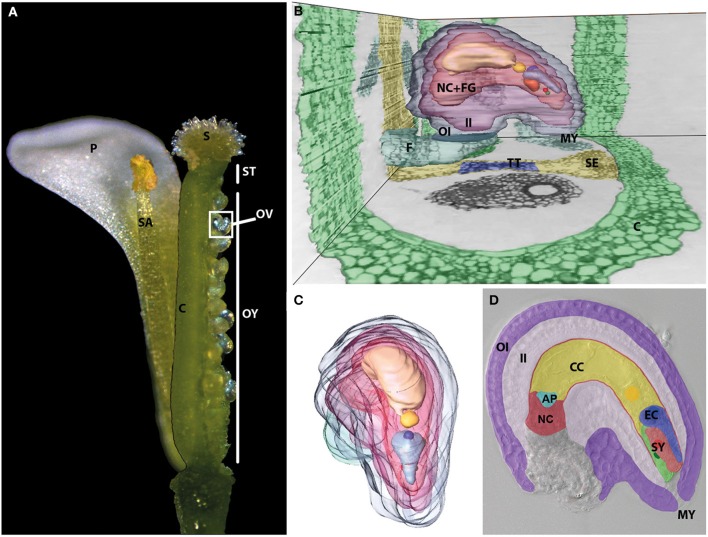
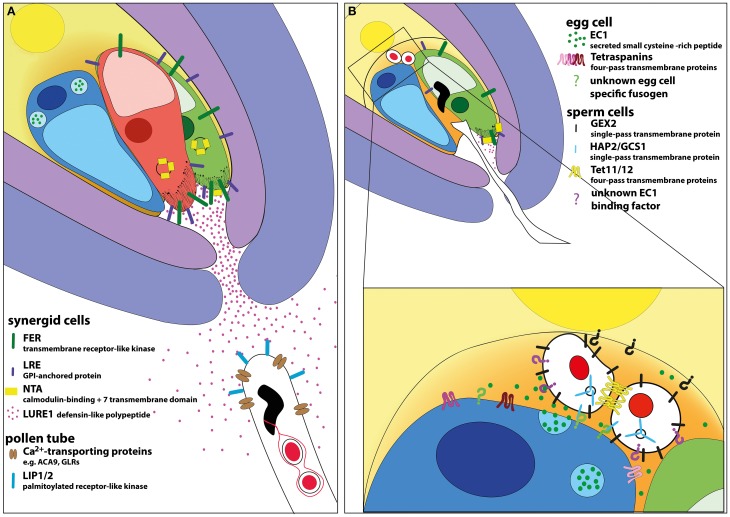
THE LAUNCH OF SEED DEVELOPMENT IN FLOWERING PLANTS (ANGIOSPERMS) IS INITIATED BY THE PROCESS OF DOUBLE FERTILIZATION: two male gametes (sperm cells) fuse with two female gametes (egg and central cell) to form the precursor cells of the two major seed components, the embryo and endosperm, respectively. The immobile sperm cells are delivered by the pollen tube toward the ovule harboring the female gametophyte by species-specific pollen tube guidance and attraction mechanisms. After pollen tube burst inside the female gametophyte, the two sperm cells fuse with the egg and central cell initiating seed development. The fertilized central cell forms the endosperm while the fertilized egg cell, the zygote, will form the actual embryo and suspensor. The latter structure connects the embryo with the sporophytic maternal tissues of the developing seed. The underlying mechanisms of double fertilization are tightly regulated to ensure delivery of functional sperm cells and the formation of both, a functional zygote and endosperm. In this review we will discuss the current state of knowledge about the processes of directed pollen tube growth and its communication with the synergid cells resulting in pollen tube burst, the interaction of the four gametes leading to cell fusion and finally discuss mechanisms how flowering plants prevent multiple sperm cell entry (polyspermy) to maximize their reproductive success.
Keywords: cell fusion; fertilization; gamete interaction; ovule; pollen tube; polyspermy; signaling.
Figures
References
Publication types
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials