

Cell entry by a novel European filovirus requires host endosomal cysteine proteases and Niemann-Pick C1
- PMID: 25310500
- PMCID: PMC4252868
- DOI: 10.1016/j.virol.2014.08.019
Cell entry by a novel European filovirus requires host endosomal cysteine proteases and Niemann-Pick C1
Abstract
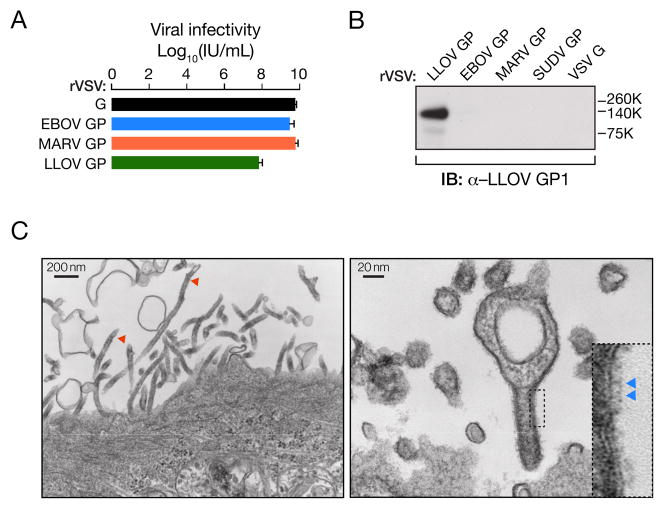
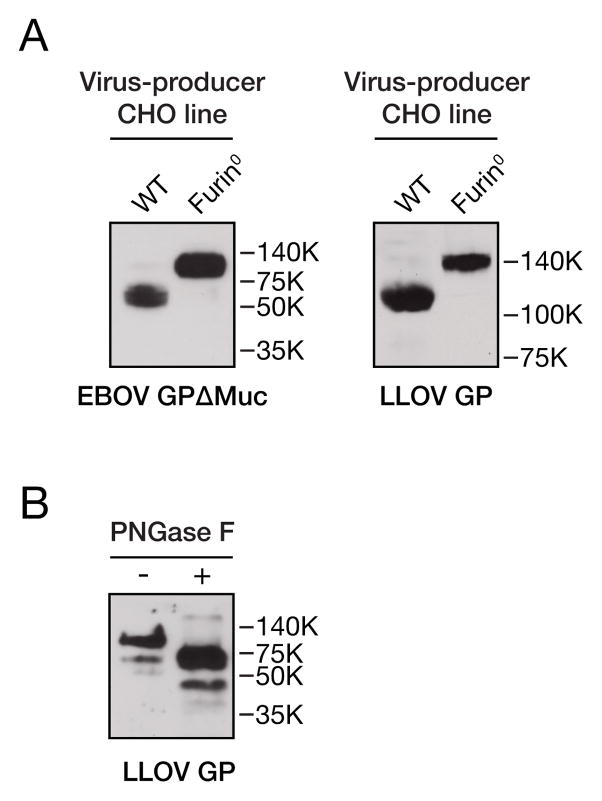
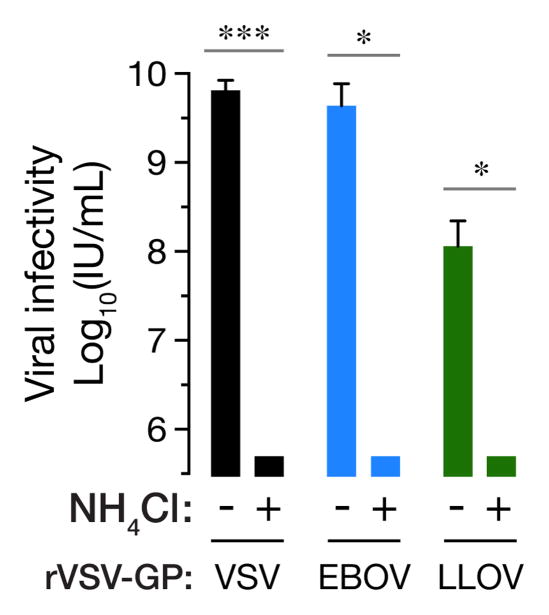
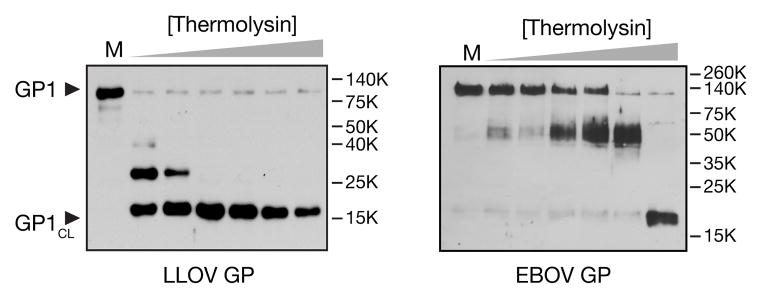
Lloviu virus (LLOV), a phylogenetically divergent filovirus, is the proposed etiologic agent of die-offs of Schreibers's long-fingered bats (Miniopterus schreibersii) in western Europe. Studies of LLOV remain limited because the infectious agent has not yet been isolated. Here, we generated a recombinant vesicular stomatitis virus expressing the LLOV spike glycoprotein (GP) and used it to show that LLOV GP resembles other filovirus GP proteins in structure and function. LLOV GP must be cleaved by endosomal cysteine proteases during entry, but is much more protease-sensitive than EBOV GP. The EBOV/MARV receptor, Niemann-Pick C1 (NPC1), is also required for LLOV entry, and its second luminal domain is recognized with high affinity by a cleaved form of LLOV GP, suggesting that receptor binding would not impose a barrier to LLOV infection of humans and non-human primates. The use of NPC1 as an intracellular entry receptor may be a universal property of filoviruses.
Keywords: Cuevavirus; Ebola; Endosomal cysteine proteases; Filoviridae; Filovirus; Lloviu virus; NPC1; Niemann–Pick C1; Viral entry; Viral glycoprotein; Viral membrane fusion; Viral receptor.
Copyright © 2014 Elsevier Inc. All rights reserved.
Figures
References
-
- Amman BR, Carroll SA, Reed ZD, Sealy TK, Balinandi S, Swanepoel R, Kemp A, Erickson BR, Comer JA, Campbell S, Cannon DL, Khristova ML, Atimnedi P, Paddock CD, Kent Crockett RJ, Flietstra TD, Warfield KL, Unfer R, Katongole-Mbidde E, Downing R, Tappero JW, Zaki SR, Rollin PE, Ksiazek TG, Nichol ST, Towner JS. Seasonal Pulses of Marburg Virus Circulation in Juvenile Rousettus aegyptiacus Bats Coincide with Periods of Increased Risk of Human Infection. PLoS Pathog. 2012;8:e1002877. doi: 10.1371/journal.ppat.1002877. - DOI - PMC - PubMed
-
- Barrette RW, Metwally SA, Rowland JM, Xu L, Zaki SR, Nichol ST, Rollin PE, Towner JS, Shieh WJ, Batten B, Sealy TK, Carrillo C, Moran KE, Bracht AJ, Mayr GA, Sirios-Cruz M, Catbagan DP, Lautner EA, Ksiazek TG, White WR, McIntosh MT. Discovery of swine as a host for the Reston ebolavirus. Science. 2009;325:204–206. doi: 10.1126/science.1172705. - DOI - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
