

Capsid expansion mechanism of bacteriophage T7 revealed by multistate atomic models derived from cryo-EM reconstructions
- PMID: 25313071
- PMCID: PMC4217468
- DOI: 10.1073/pnas.1407020111
Capsid expansion mechanism of bacteriophage T7 revealed by multistate atomic models derived from cryo-EM reconstructions
Abstract
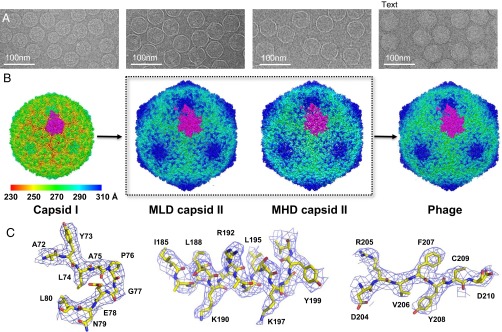
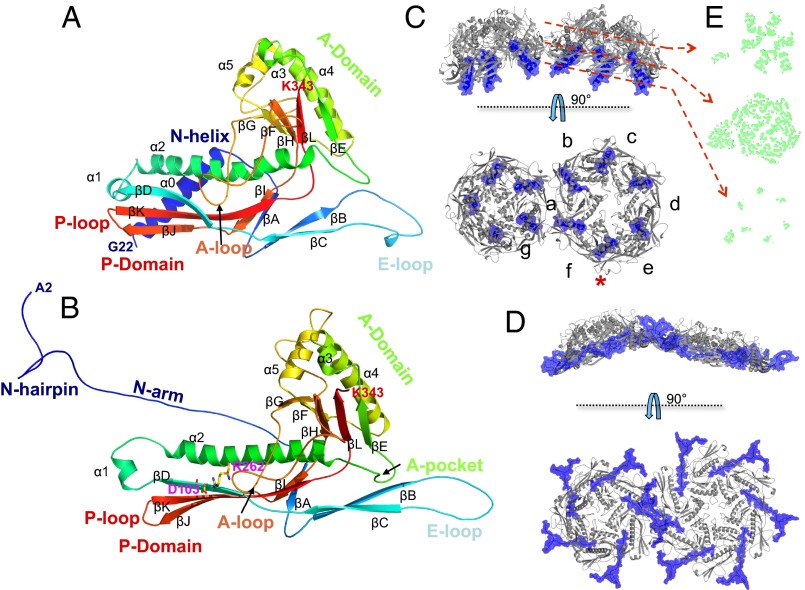
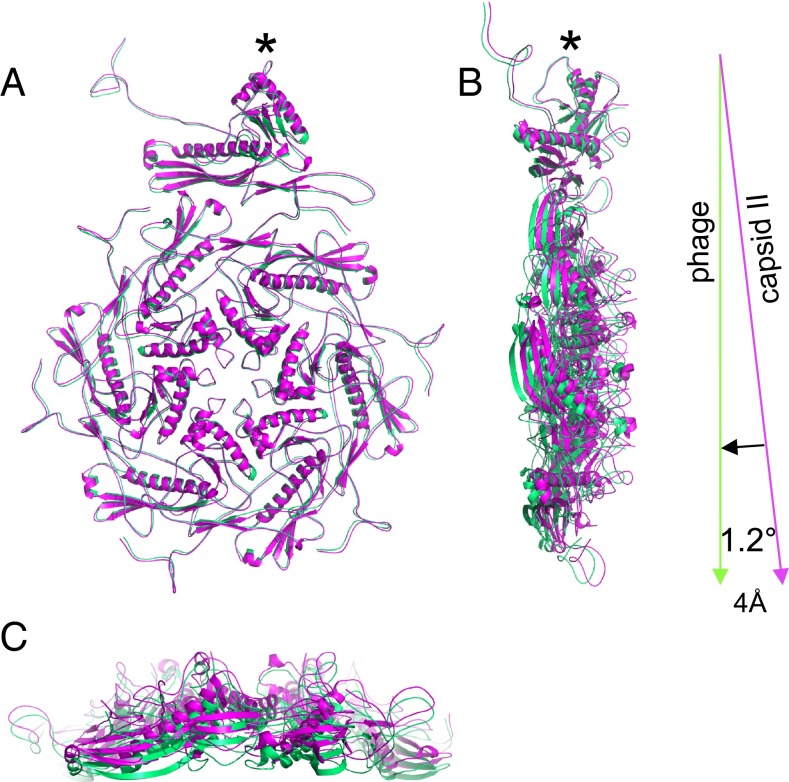
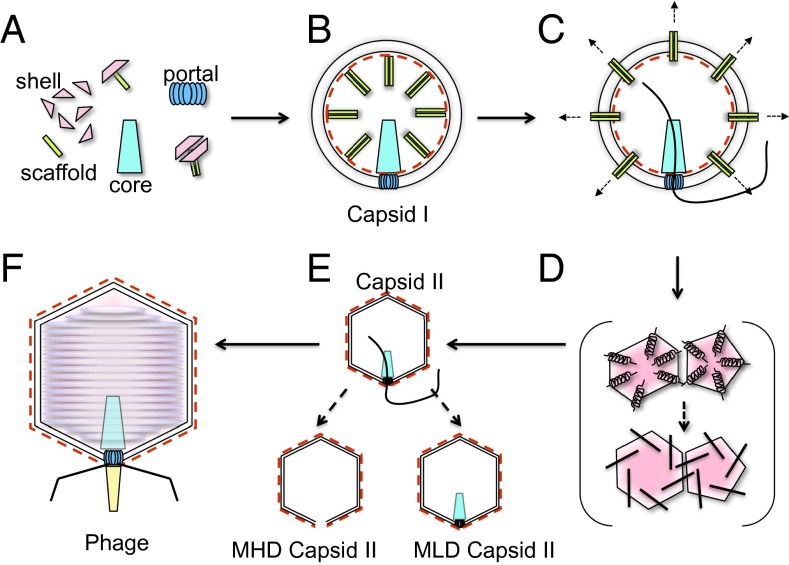
Many dsDNA viruses first assemble a DNA-free procapsid, using a scaffolding protein-dependent process. The procapsid, then, undergoes dramatic conformational maturation while packaging DNA. For bacteriophage T7 we report the following four single-particle cryo-EM 3D reconstructions and the derived atomic models: procapsid (4.6-Å resolution), an early-stage DNA packaging intermediate (3.5 Å), a later-stage packaging intermediate (6.6 Å), and the final infectious phage (3.6 Å). In the procapsid, the N terminus of the major capsid protein, gp10, has a six-turn helix at the inner surface of the shell, where each skewed hexamer of gp10 interacts with two scaffolding proteins. With the exit of scaffolding proteins during maturation the gp10 N-terminal helix unfolds and swings through the capsid shell to the outer surface. The refolded N-terminal region has a hairpin that forms a novel noncovalent, joint-like, intercapsomeric interaction with a pocket formed during shell expansion. These large conformational changes also result in a new noncovalent, intracapsomeric topological linking. Both interactions further stabilize the capsids by interlocking all pentameric and hexameric capsomeres in both DNA packaging intermediate and phage. Although the final phage shell has nearly identical structure to the shell of the DNA-free intermediate, surprisingly we found that the icosahedral faces of the phage are slightly (∼4 Å) contracted relative to the faces of the intermediate, despite the internal pressure from the densely packaged DNA genome. These structures provide a basis for understanding the capsid maturation process during DNA packaging that is essential for large numbers of dsDNA viruses.
Keywords: DNA packaging intermediates; bacteriophage T7 maturation; noncovalent topological linking; procapsid; single-particle cryo-EM.
Conflict of interest statement
The authors declare no conflict of interest.
Figures


References
-
- Jiang W, et al. Coat protein fold and maturation transition of bacteriophage P22 seen at subnanometer resolutions. Nat Struct Biol. 2003;10(2):131–135. - PubMed
-
- Cerritelli ME, Conway JF, Cheng N, Trus BL, Steven AC. Molecular mechanisms in bacteriophage T7 procapsid assembly, maturation, and DNA containment. Adv Protein Chem. 2003;64:301–323. - PubMed
-
- Conway JF, et al. Virus maturation involving large subunit rotations and local refolding. Science. 2001;292(5517):744–748. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
