

Bacterially produced Pt-GFP as ratiometric dual-excitation sensor for in planta mapping of leaf apoplastic pH in intact Avena sativa and Vicia faba
- PMID: 25313311
- PMCID: PMC4194361
- DOI: 10.1186/1746-4811-10-31
Bacterially produced Pt-GFP as ratiometric dual-excitation sensor for in planta mapping of leaf apoplastic pH in intact Avena sativa and Vicia faba
Abstract
Background: Ratiometric analysis with H(+)-sensitive fluorescent sensors is a suitable approach for monitoring apoplastic pH dynamics. For the acidic range, the acidotropic dual-excitation dye Oregon Green 488 is an excellent pH sensor. Long lasting (hours) recordings of apoplastic pH in the near neutral range, however, are more problematic because suitable pH indicators that combine a good pH responsiveness at a near neutral pH with a high photostability are lacking. The fluorescent pH reporter protein from Ptilosarcus gurneyi (Pt-GFP) comprises both properties. But, as a genetically encoded indicator and expressed by the plant itself, it can be used almost exclusively in readily transformed plants. In this study we present a novel approach and use purified recombinant indicators for measuring ion concentrations in the apoplast of crop plants such as Vicia faba L. and Avena sativa L.
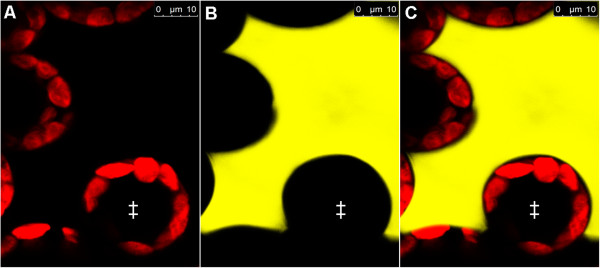
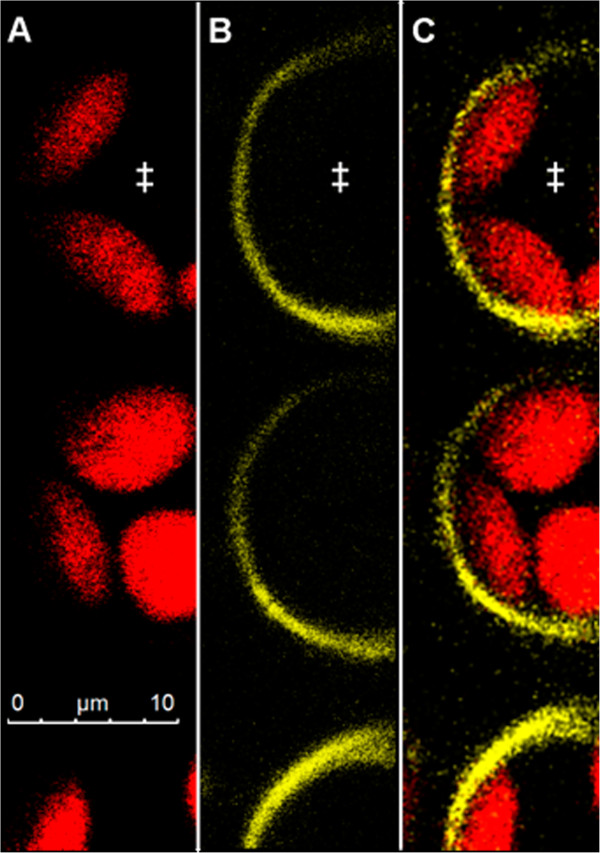
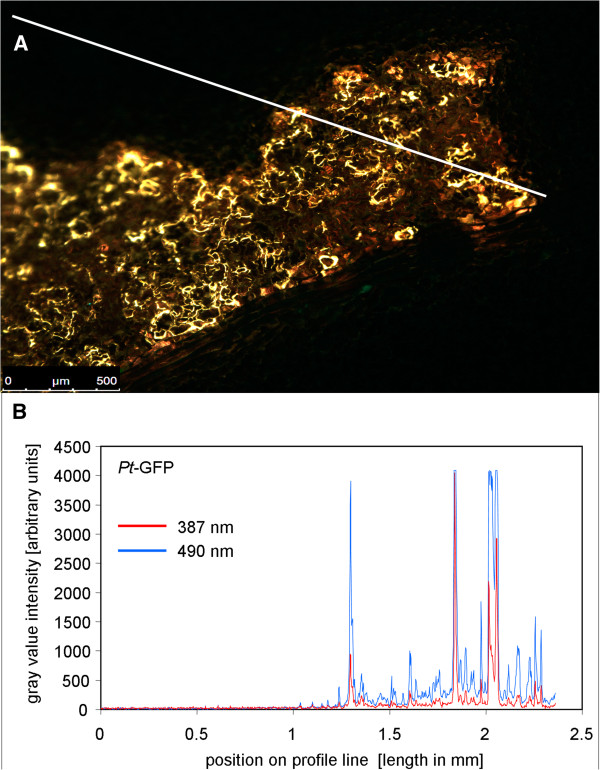
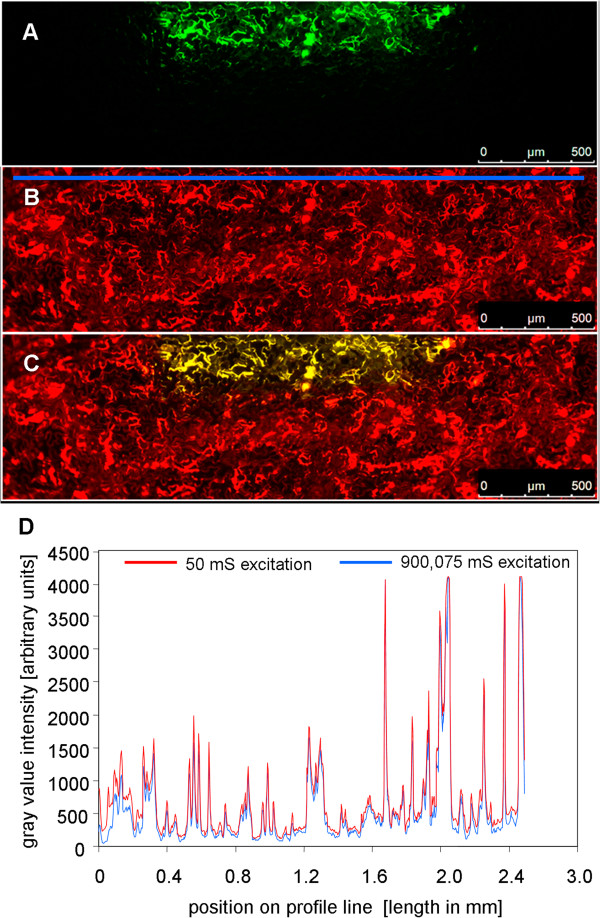
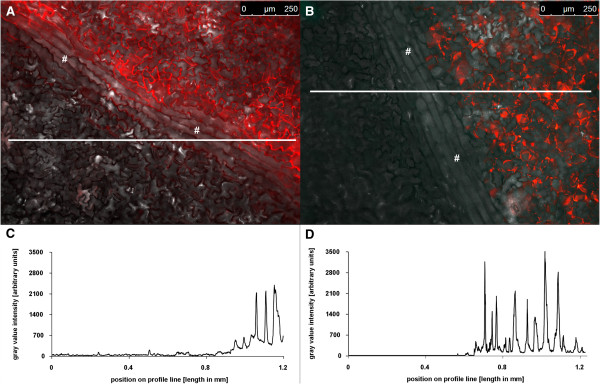
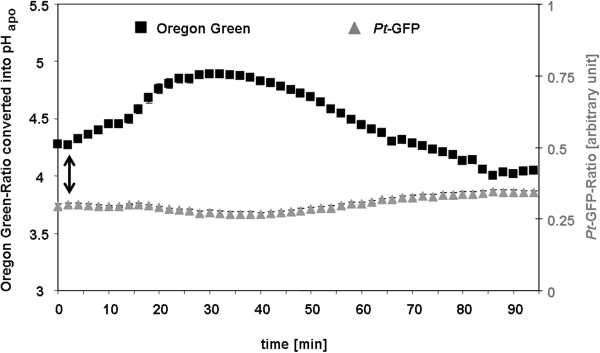
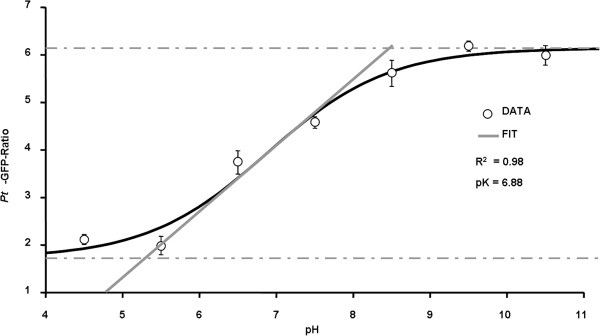
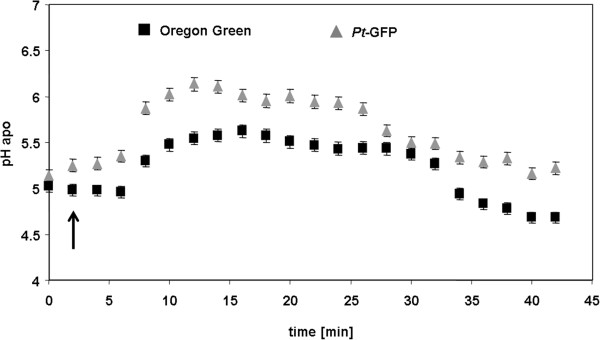
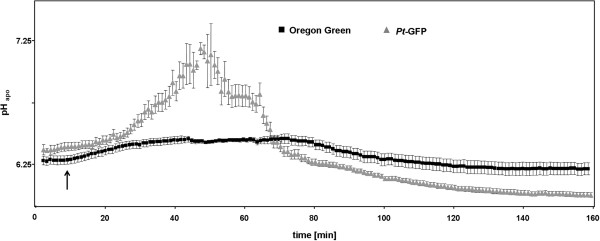
Results: Pt-GFP was purified using a bacterial expression system and subsequently loaded through stomata into the leaf apoplast of intact plants. Imaging verified the apoplastic localization of Pt-GFP and excluded its presence in the symplast. The pH-dependent emission signal stood out clearly from the background. PtGFP is highly photostable, allowing ratiometric measurements over hours. By using this approach, a chloride-induced alkalinizations of the apoplast was demonstrated for the first in oat.
Conclusions: Pt-GFP appears to be an excellent sensor for the quantification of leaf apoplastic pH in the neutral range. The presented approach encourages to also use other genetically encoded biosensors for spatiotemporal mapping of apoplastic ion dynamics.
Keywords: Apoplast; GFP; Genetically encoded biosensor; Nitrogen forms; Plant bioimaging; Ptilosarcus gurneyi; Salinity; Signaling; Stress; Three-channel ratio imaging; pH.
Figures

References
-
- Lemoine R, Delrot S. Proton-motive-force-driven sucrose uptake in sugar beet plasma membrane vesicles. FEBS Lett. 1989;249:129. doi: 10.1016/0014-5793(89)80030-4. - DOI
-
- Bush DR. Proton-coupled sugar and amino acid transporters in plants. Annu Rev Plant Physiol Plant Mol Biol. 1993;44:513–542. doi: 10.1146/annurev.pp.44.060193.002501. - DOI
LinkOut - more resources
Full Text Sources
Other Literature Sources
