

Asynchronous remodeling is a driver of failed regeneration in Duchenne muscular dystrophy
- PMID: 25313409
- PMCID: PMC4195829
- DOI: 10.1083/jcb.201402079
Asynchronous remodeling is a driver of failed regeneration in Duchenne muscular dystrophy
Abstract
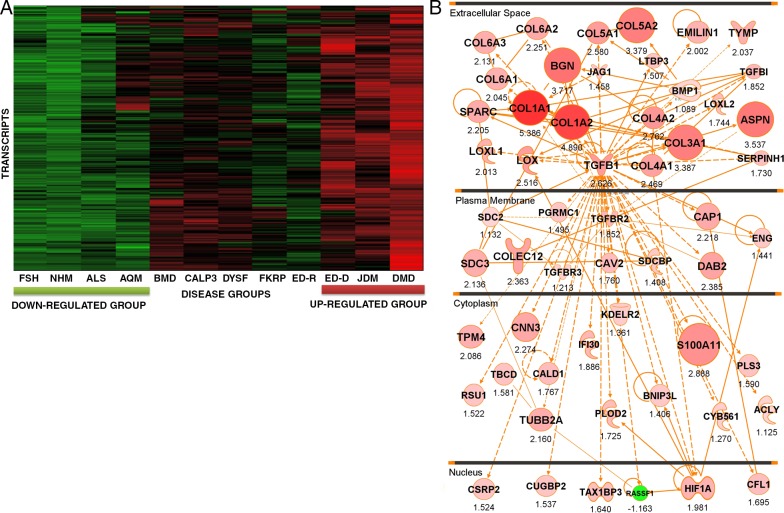
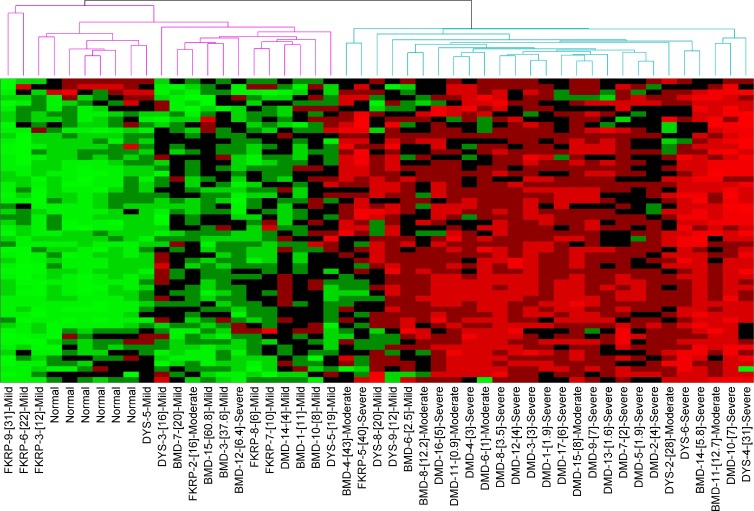
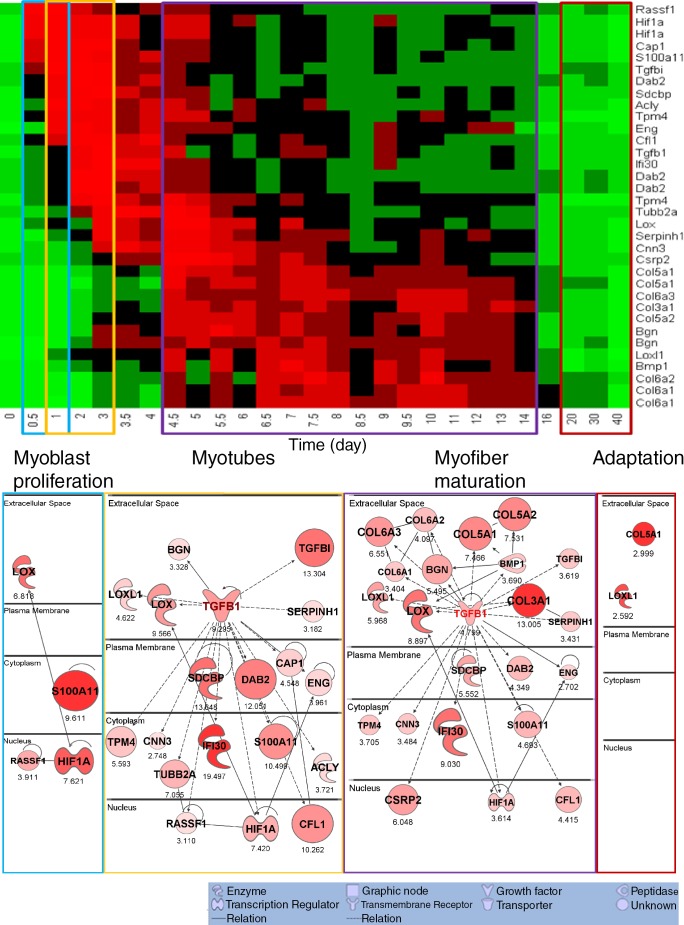
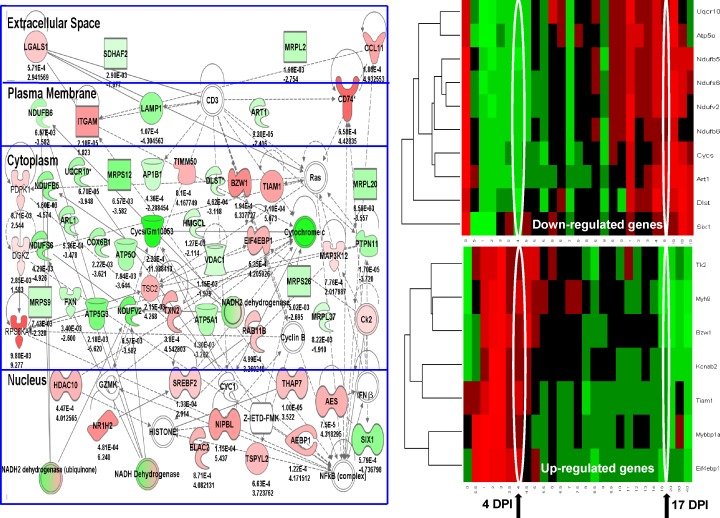
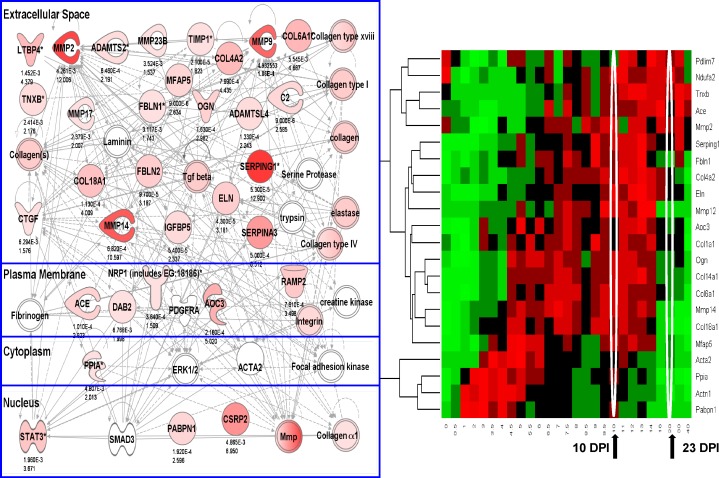
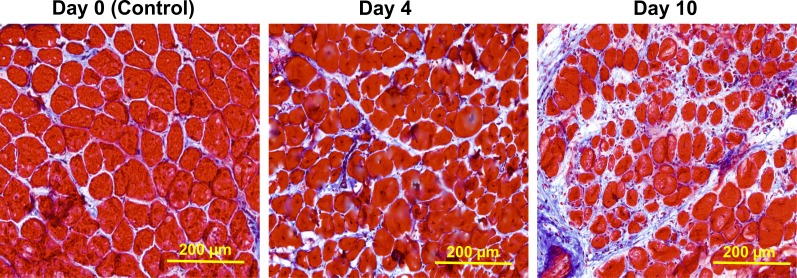
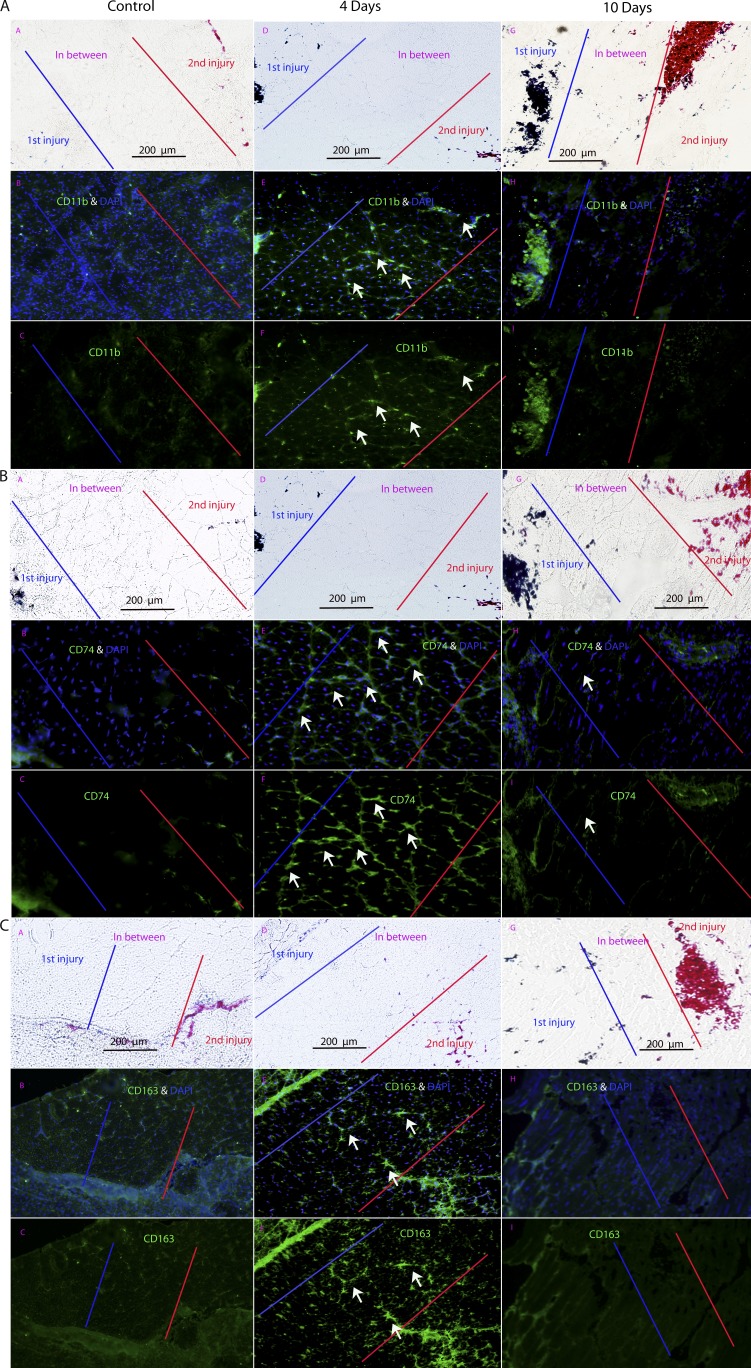
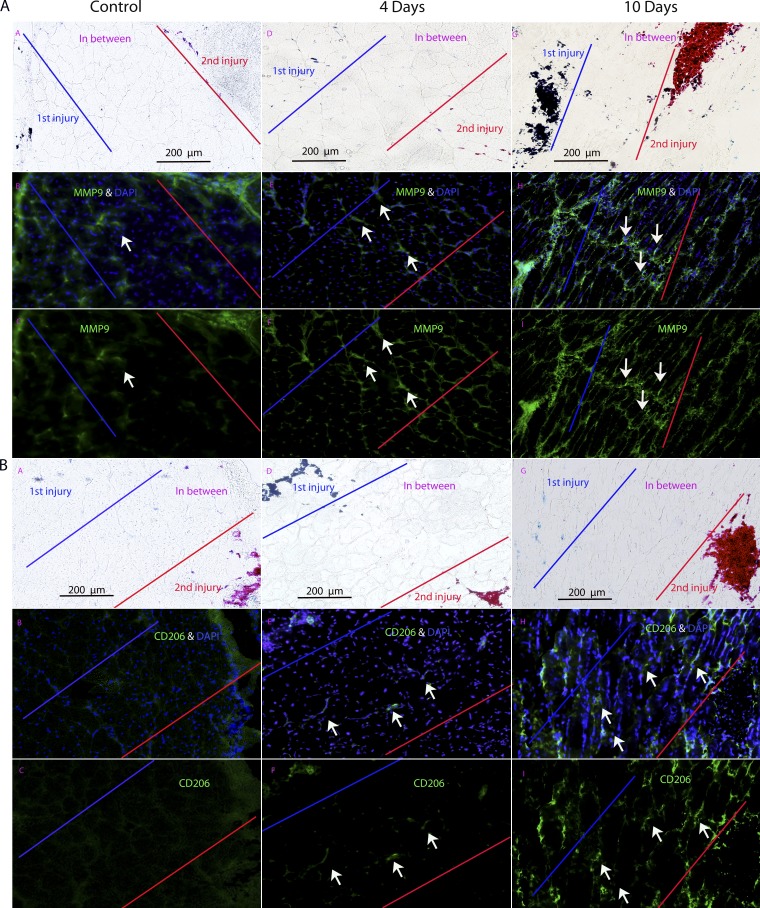
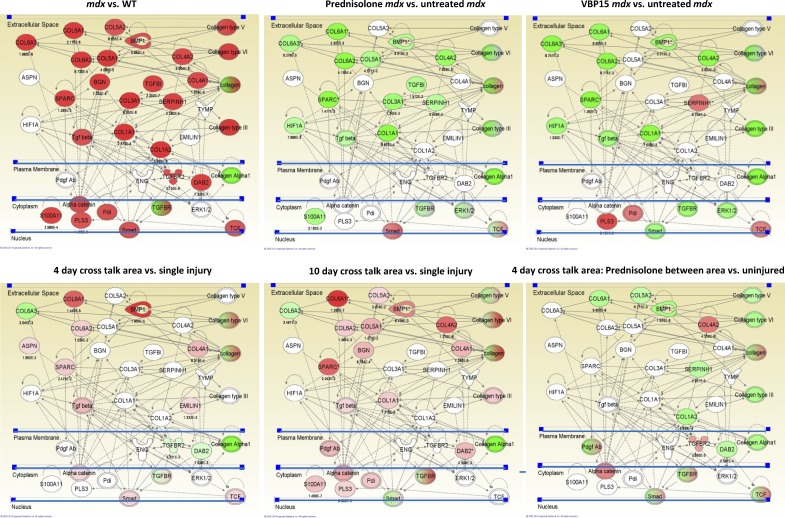
We sought to determine the mechanisms underlying failure of muscle regeneration that is observed in dystrophic muscle through hypothesis generation using muscle profiling data (human dystrophy and murine regeneration). We found that transforming growth factor β-centered networks strongly associated with pathological fibrosis and failed regeneration were also induced during normal regeneration but at distinct time points. We hypothesized that asynchronously regenerating microenvironments are an underlying driver of fibrosis and failed regeneration. We validated this hypothesis using an experimental model of focal asynchronous bouts of muscle regeneration in wild-type (WT) mice. A chronic inflammatory state and reduced mitochondrial oxidative capacity are observed in bouts separated by 4 d, whereas a chronic profibrotic state was seen in bouts separated by 10 d. Treatment of asynchronously remodeling WT muscle with either prednisone or VBP15 mitigated the molecular phenotype. Our asynchronous regeneration model for pathological fibrosis and muscle wasting in the muscular dystrophies is likely generalizable to tissue failure in chronic inflammatory states in other regenerative tissues.
© 2014 Dadgar et al.
Figures
References
-
- Aguennouz, M., Vita G.L., Messina S., Cama A., Lanzano N., Ciranni A., Rodolico C., Di Giorgio R.M., and Vita G.. 2011. Telomere shortening is associated to TRF1 and PARP1 overexpression in Duchenne muscular dystrophy. Neurobiol. Aging. 32:2190–2197 10.1016/j.neurobiolaging.2010.01.008 - DOI - PubMed
-
- Alcala, S.E., Benton A.S., Watson A.M., Kureshi S., Reeves E.M., Damsker J., Wang Z., Nagaraju K., Anderson J., Williams A.M., et al. 2014. Mitotic asynchrony induces transforming growth factor-β1 secretion from airway epithelium. Am. J. Respir. Cell Mol. Biol. 51:363–369 10.1165/rcmb.2013-0396OC - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
