

A conserved endoplasmic reticulum membrane protein complex (EMC) facilitates phospholipid transfer from the ER to mitochondria
- PMID: 25313861
- PMCID: PMC4196738
- DOI: 10.1371/journal.pbio.1001969
A conserved endoplasmic reticulum membrane protein complex (EMC) facilitates phospholipid transfer from the ER to mitochondria
Abstract
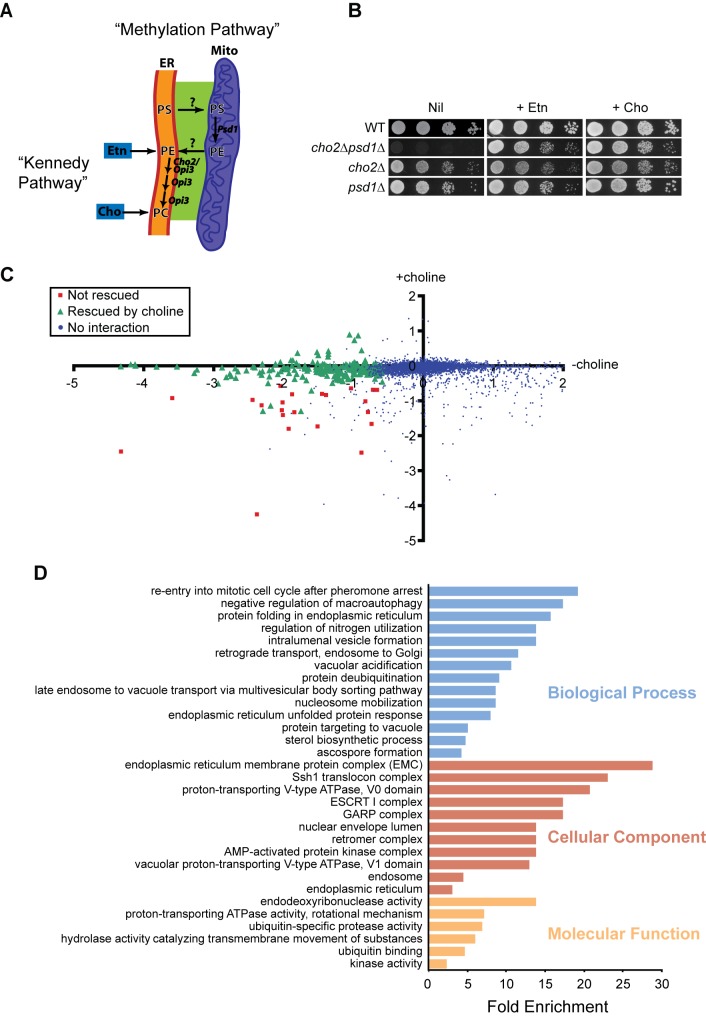
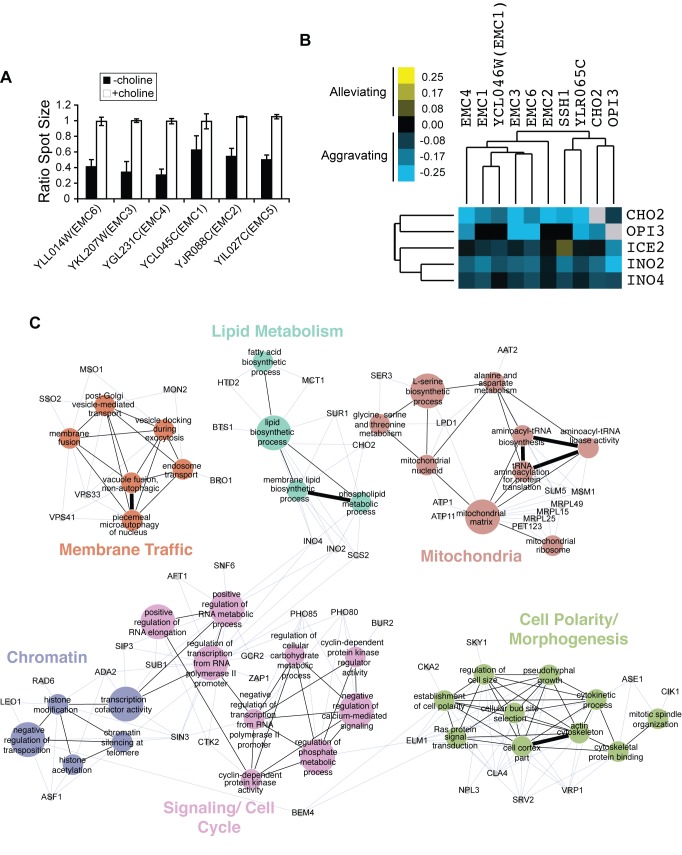
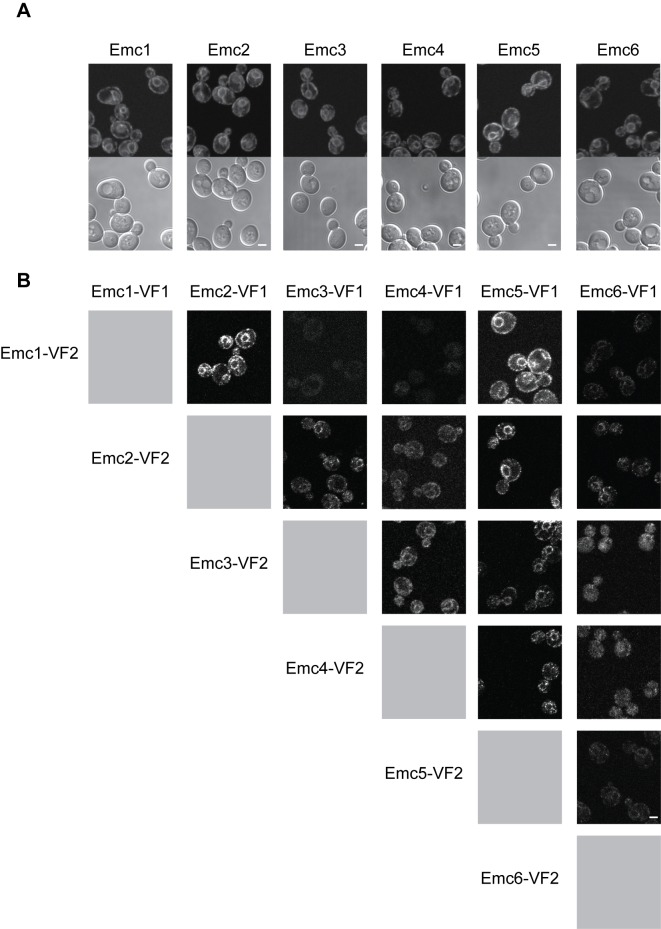
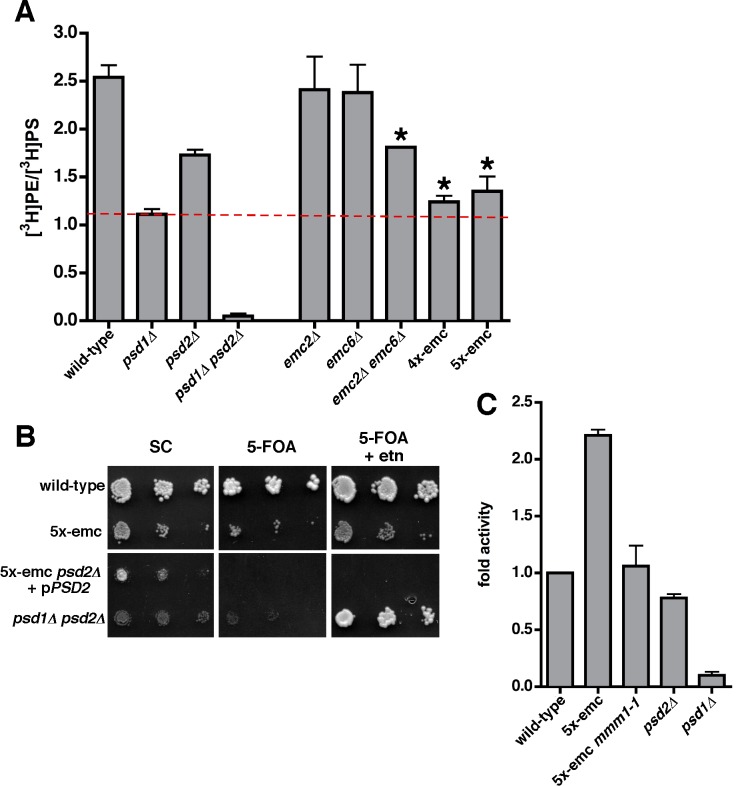
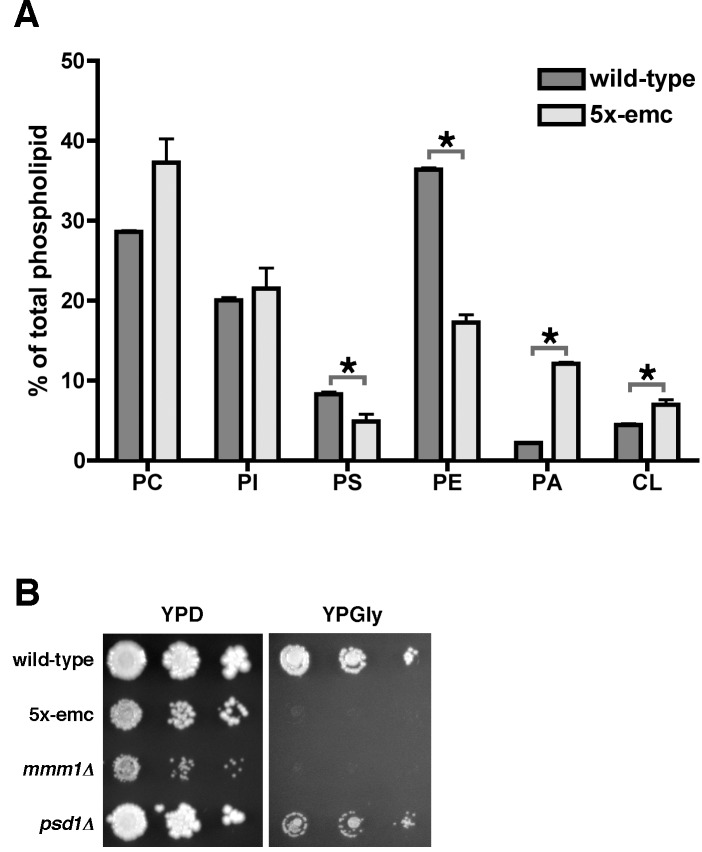
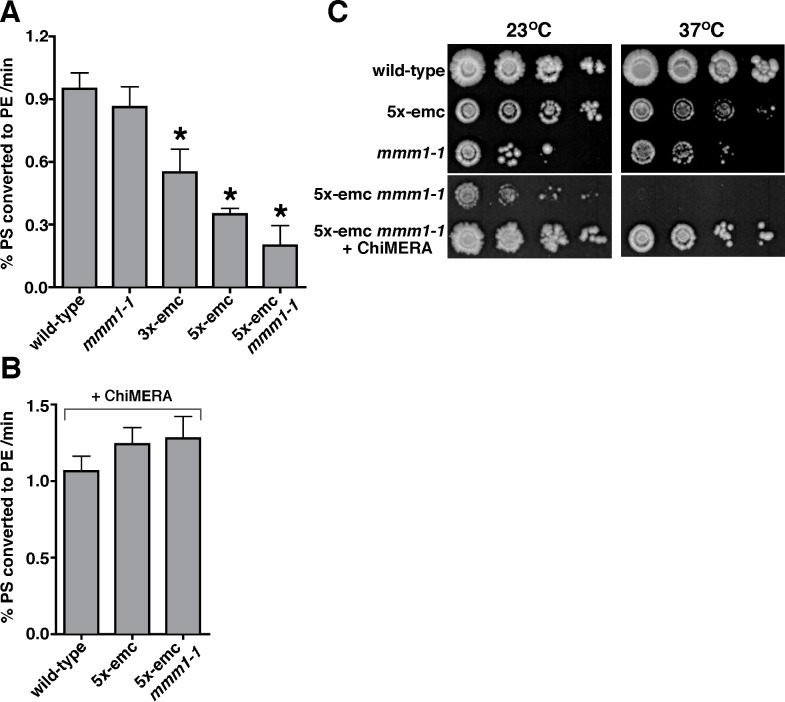
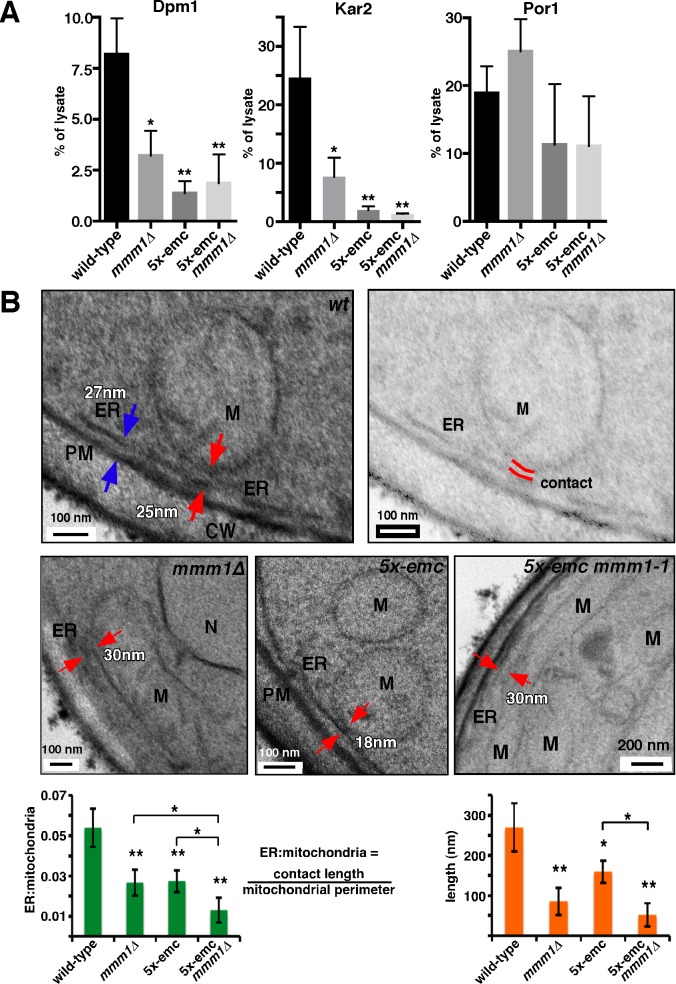
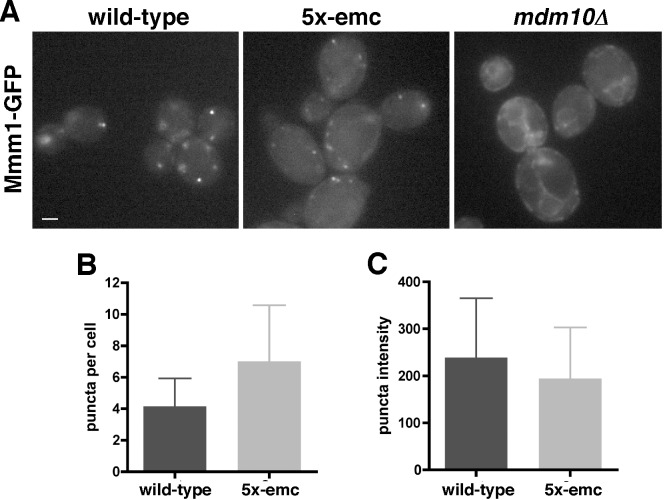
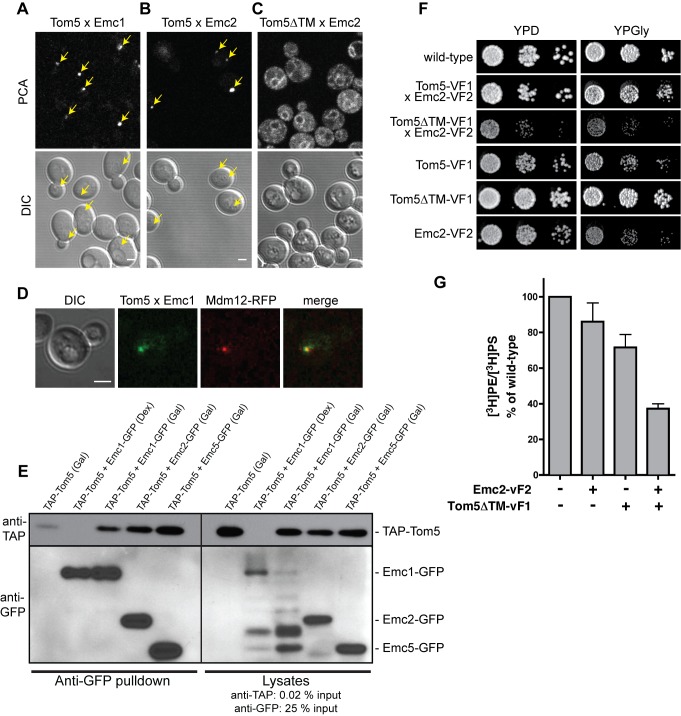
Mitochondrial membrane biogenesis and lipid metabolism require phospholipid transfer from the endoplasmic reticulum (ER) to mitochondria. Transfer is thought to occur at regions of close contact of these organelles and to be nonvesicular, but the mechanism is not known. Here we used a novel genetic screen in S. cerevisiae to identify mutants with defects in lipid exchange between the ER and mitochondria. We show that a strain missing multiple components of the conserved ER membrane protein complex (EMC) has decreased phosphatidylserine (PS) transfer from the ER to mitochondria. Mitochondria from this strain have significantly reduced levels of PS and its derivative phosphatidylethanolamine (PE). Cells lacking EMC proteins and the ER-mitochondria tethering complex called ERMES (the ER-mitochondria encounter structure) are inviable, suggesting that the EMC also functions as a tether. These defects are corrected by expression of an engineered ER-mitochondrial tethering protein that artificially tethers the ER to mitochondria. EMC mutants have a significant reduction in the amount of ER tethered to mitochondria even though ERMES remained intact in these mutants, suggesting that the EMC performs an additional tethering function to ERMES. We find that all Emc proteins interact with the mitochondrial translocase of the outer membrane (TOM) complex protein Tom5 and this interaction is important for PS transfer and cell growth, suggesting that the EMC forms a tether by associating with the TOM complex. Together, our findings support that the EMC tethers ER to mitochondria, which is required for phospholipid synthesis and cell growth.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
-
- Elbaz Y, Schuldiner M (2011) Staying in touch: the molecular era of organelle contact sites. Trends Biochem Sci 36: 616–623. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
