

Anatomical localization of progenitor cells in human breast tissue reveals enrichment of uncommitted cells within immature lobules
- PMID: 25315014
- PMCID: PMC4303132
- DOI: 10.1186/s13058-014-0453-3
Anatomical localization of progenitor cells in human breast tissue reveals enrichment of uncommitted cells within immature lobules
Abstract
Introduction: Lineage tracing studies in mice have revealed the localization and existence of lineage-restricted mammary epithelial progenitor cells that functionally contribute to expansive growth during puberty and differentiation during pregnancy. However, extensive anatomical differences between mouse and human mammary tissues preclude the direct translation of rodent findings to the human breast. Therefore, here we characterize the mammary progenitor cell hierarchy and identify the anatomic location of progenitor cells within human breast tissues.
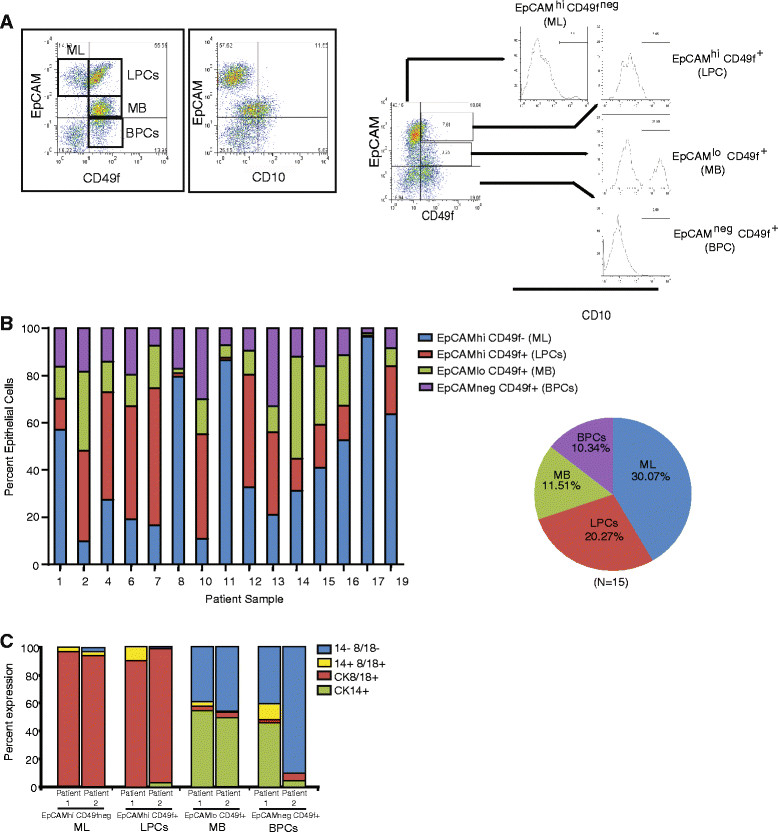
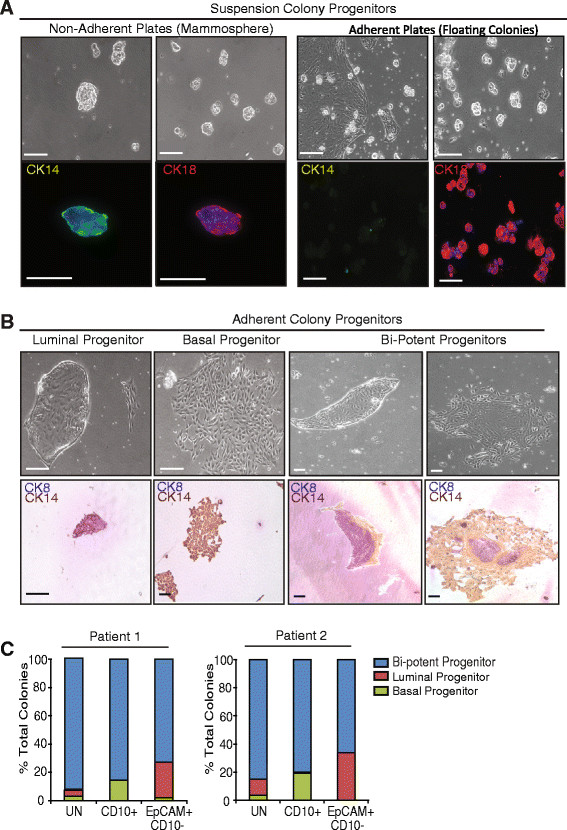
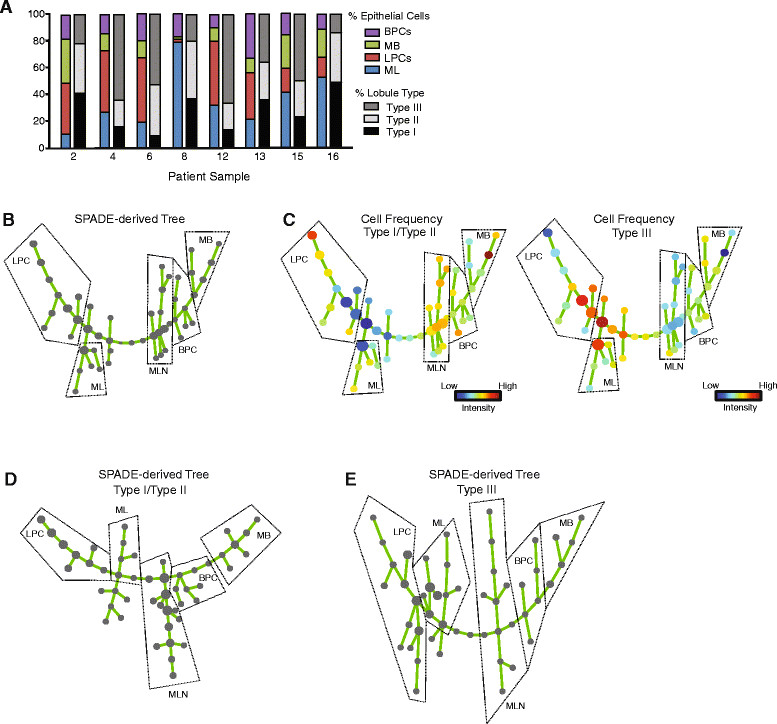
Methods: Mammary epithelial cells (MECs) were isolated from disease-free reduction mammoplasty tissues and assayed for stem/progenitor activity in vitro and in vivo. MECs were sorted and evaluated for growth on collagen and expression of lineages markers. Breast lobules were microdissected and individually characterized based on lineage markers and steroid receptor expression to identify the anatomic location of progenitor cells. Spanning-tree progression analysis of density-normalized events (SPADE) was used to identify the cellular hierarchy of MECs within lobules from high-dimensional cytometry data.
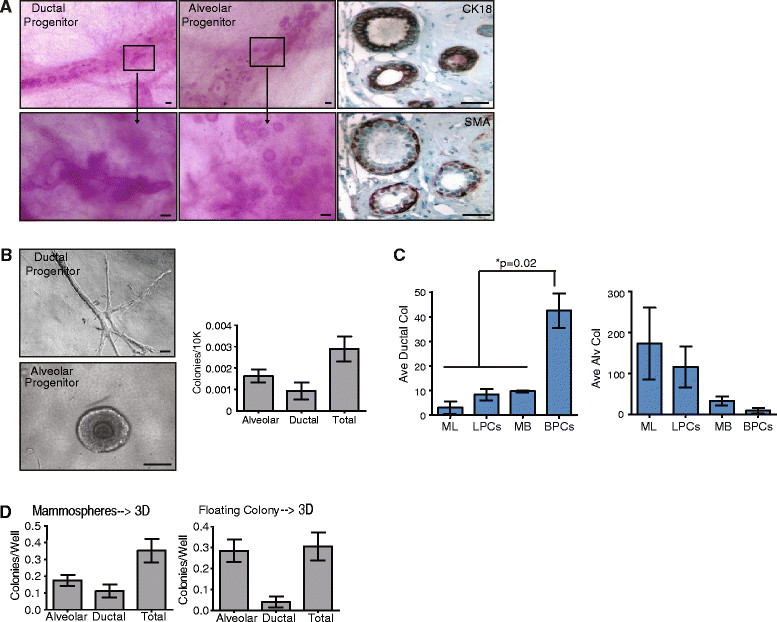
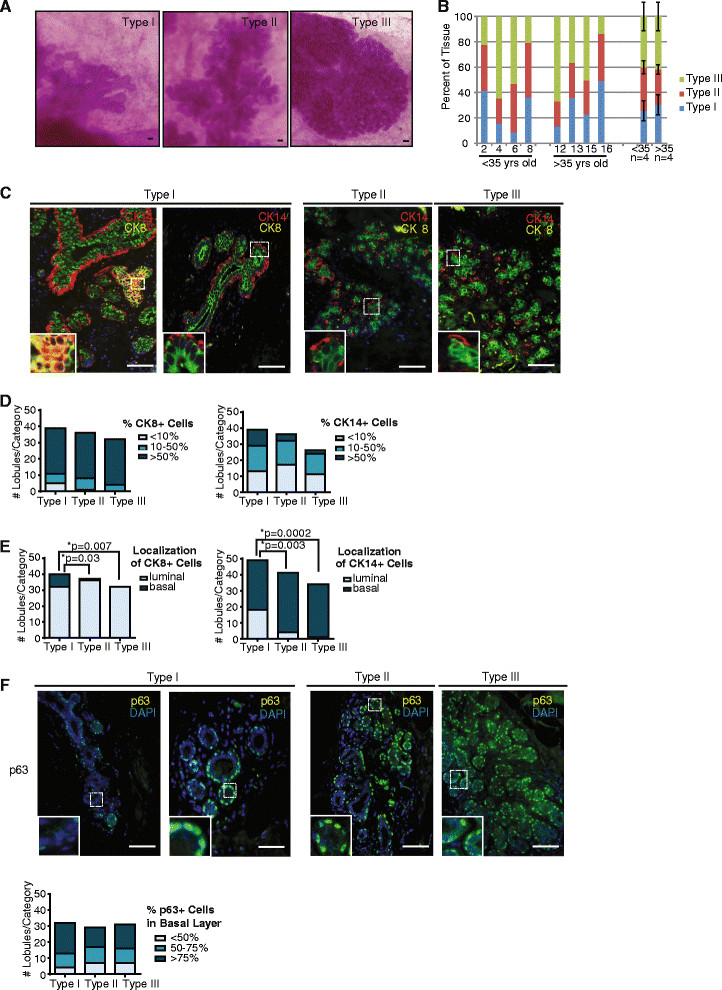
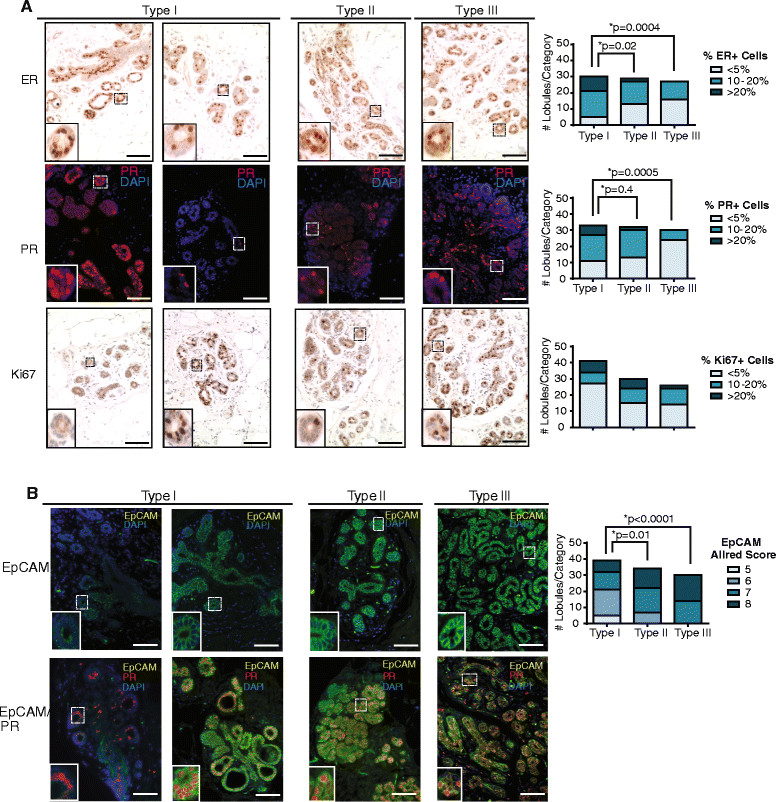
Results: Integrating multiple assays for progenitor activity, we identified the presence of luminal alveolar and basal ductal progenitors. Further, we show that Type I lobules of the human breast were the least mature, demonstrating an unrestricted pattern of expression of luminal and basal lineage markers. Consistent with this, SPADE analysis revealed that immature lobules were enriched for basal progenitor cells, while mature lobules consisted of increased hierarchal complexity of cells within the luminal lineages.
Conclusions: These results reveal underlying differences in the human breast epithelial hierarchy and suggest that with increasing glandular maturity, the epithelial hierarchy also becomes more complex.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
