

Macrophage proliferation, provenance, and plasticity in macroparasite infection
- PMID: 25319331
- PMCID: PMC4324133
- DOI: 10.1111/imr.12221
Macrophage proliferation, provenance, and plasticity in macroparasite infection
Abstract
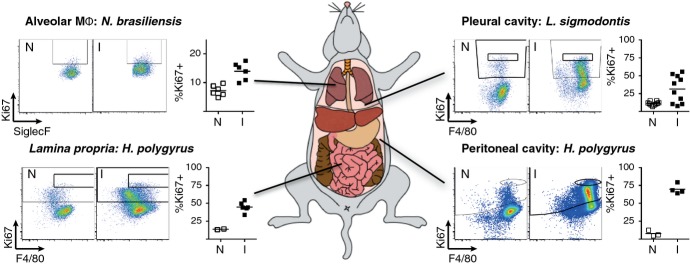
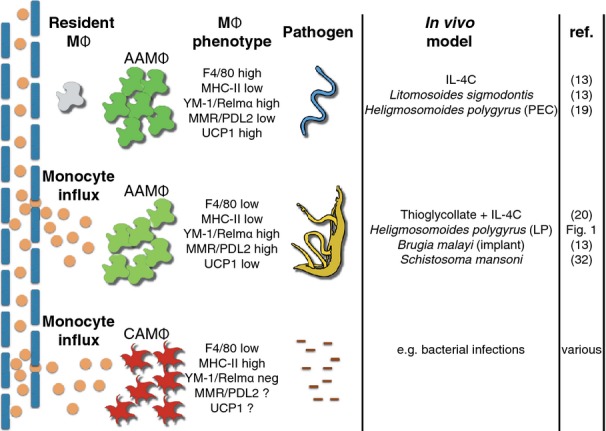
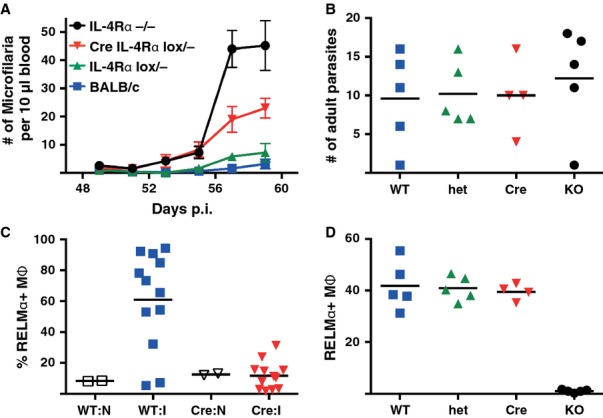
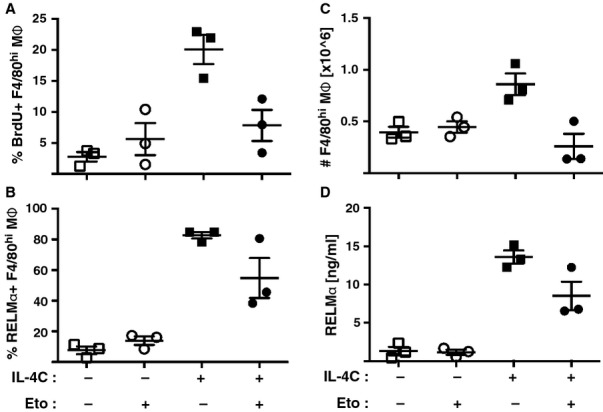
Macrophages have long been center stage in the host response to microbial infection, but only in the past 10-15 years has there been a growing appreciation for their role in helminth infection and the associated type 2 response. Through the actions of the IL-4 receptor α (IL-4Rα), type 2 cytokines result in the accumulation of macrophages with a distinctive activation phenotype. Although our knowledge of IL-4Rα-induced genes is growing rapidly, the specific functions of these macrophages have yet to be established in most disease settings. Understanding the interplay between IL-4Rα-activated macrophages and the other cellular players is confounded by the enormous transcriptional heterogeneity within the macrophage population and by their highly plastic nature. Another level of complexity is added by the new knowledge that tissue macrophages can be derived either from a resident prenatal population or from blood monocyte recruitment and that IL-4 can increase macrophage numbers through proliferative expansion. Here, we review current knowledge on the contribution of macrophages to helminth killing and wound repair, with specific attention paid to distinct cellular origins and plasticity potential.
Keywords: IL-4 receptor; helminth infection; inflammation; macrophage; monocyte; wound repair.
© 2014 The Authors. Immunological Reviews published by John Wiley & Sons Ltd.
Figures
References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
