

Conformational response to ligand binding in phosphomannomutase2: insights into inborn glycosylation disorder
- PMID: 25324542
- PMCID: PMC4263888
- DOI: 10.1074/jbc.M114.586362
Conformational response to ligand binding in phosphomannomutase2: insights into inborn glycosylation disorder
Abstract
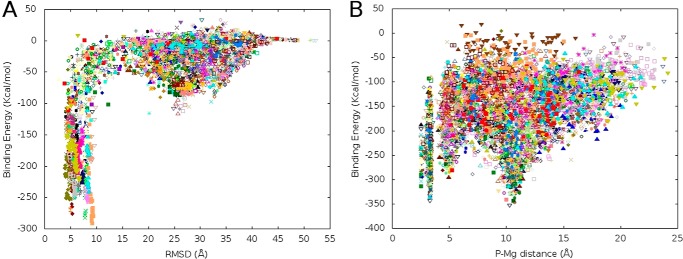
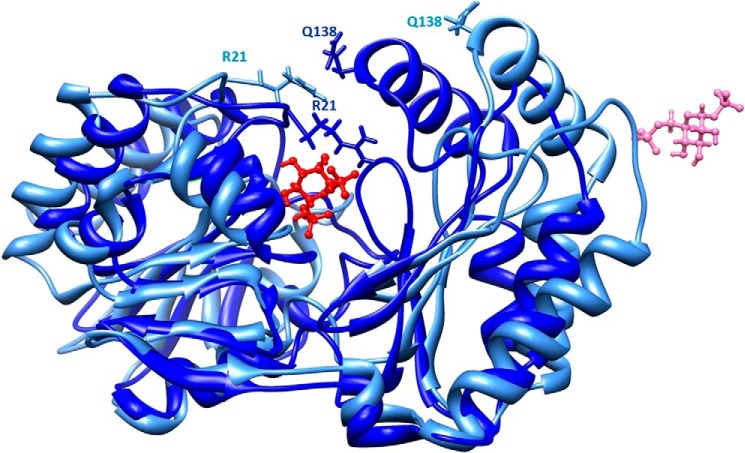
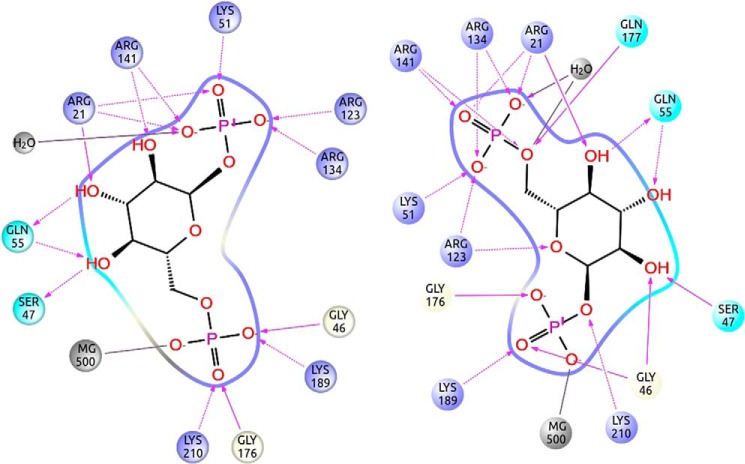
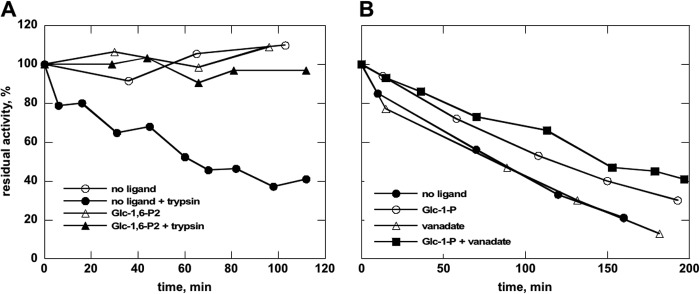
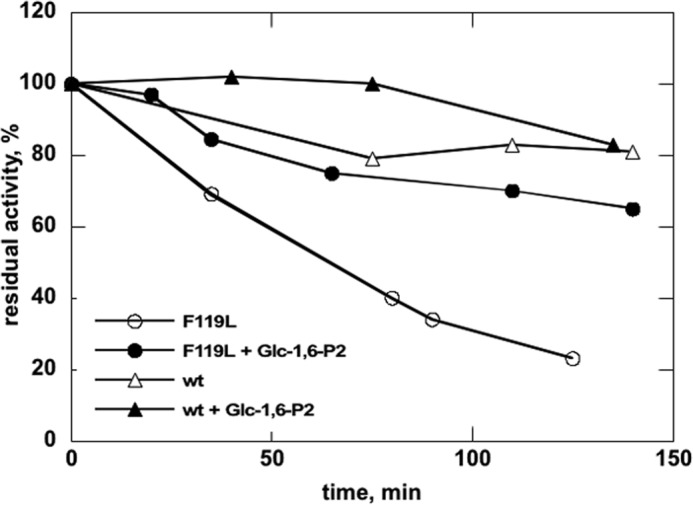
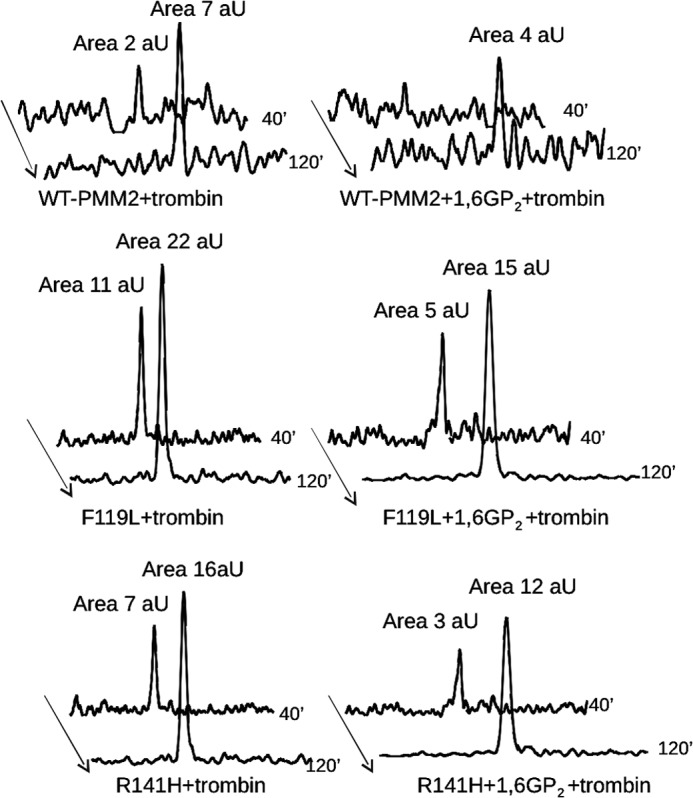
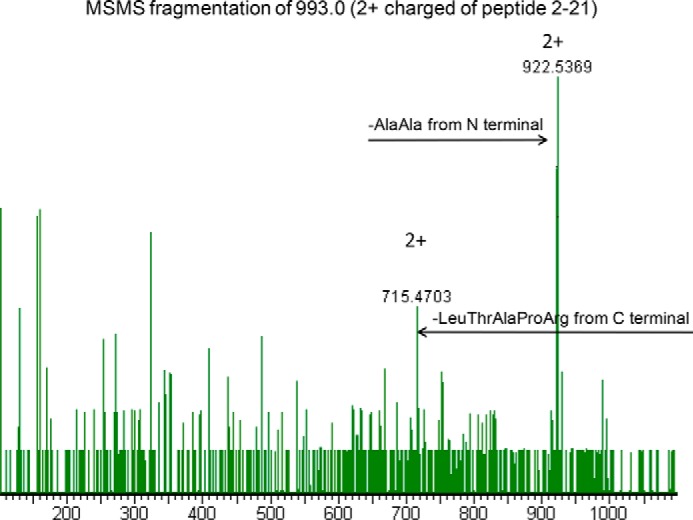
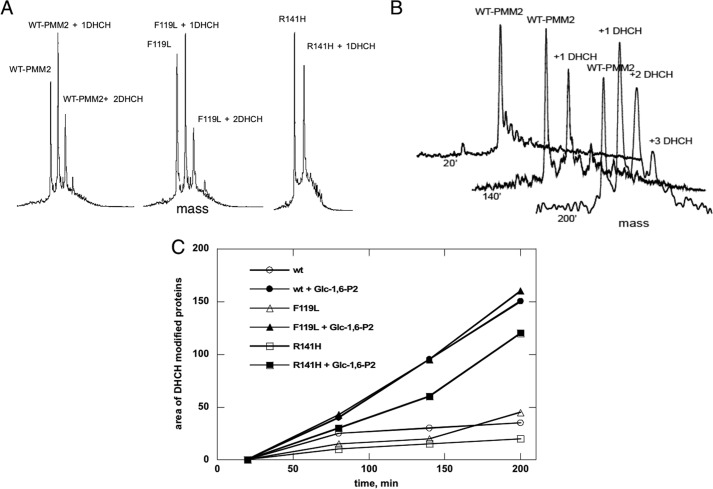
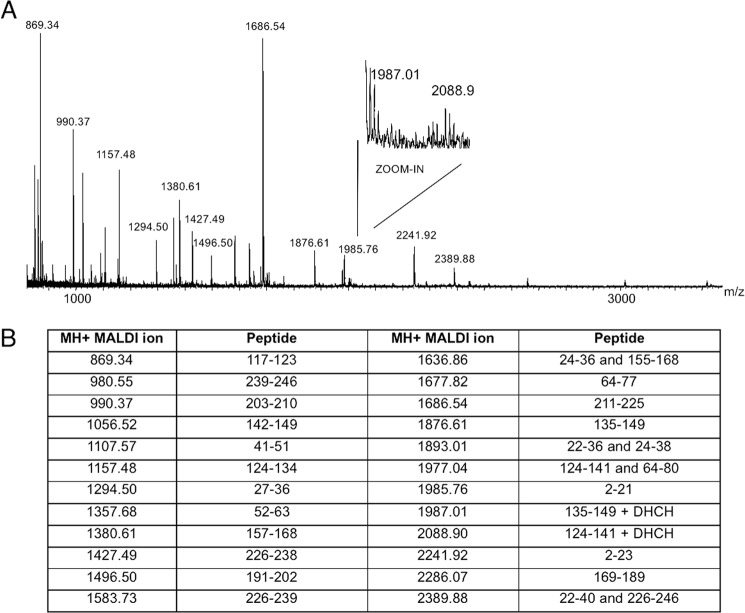
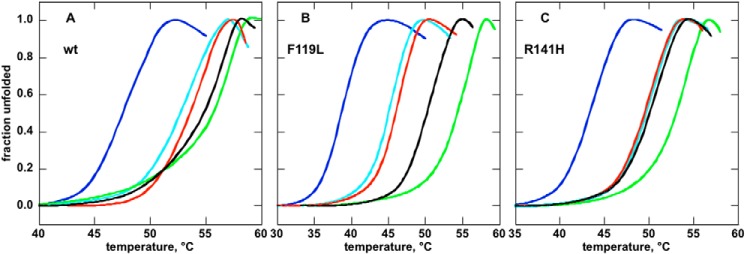
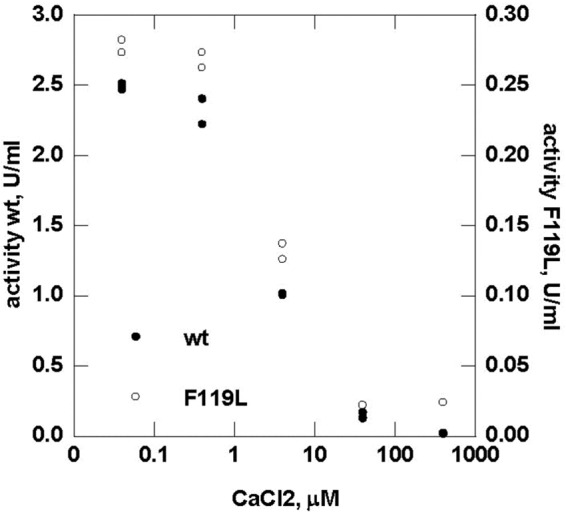
The most common glycosylation disorder is caused by mutations in the gene encoding phosphomannomutase2, producing a disease still without a cure. Phosphomannomutase2, a homodimer in which each chain is composed of two domains, requires a bisphosphate sugar (either mannose or glucose) as activator, opening a possible drug design path for therapeutic purposes. The crystal structure of human phosphomannomutase2, however, lacks bound substrate and a key active site loop. To speed up drug discovery, we present here the first structural model of a bisphosphate substrate bound to human phosphomannomutase2. Taking advantage of recent developments in all-atom simulation techniques in combination with limited and site-directed proteolysis, we demonstrated that α-glucose 1,6-bisphosphate can adopt two low energy orientations as required for catalysis. Upon ligand binding, the two domains come close, making the protein more compact, in analogy to the enzyme in the crystals from Leishmania mexicana. Moreover, proteolysis was also carried out on two common mutants, R141H and F119L. It was an unexpected finding that the mutant most frequently found in patients, R141H, although inactive, does bind α-glucose 1,6-bisphosphate and changes conformation.
Keywords: 1,6-Bisphosphate; Computer Modeling; Drug Discovery; Glycosylation; Glycosylation Inhibitor; Ligand-binding Protein; PELE; Phosphomannomutase.
© 2014 by The American Society for Biochemistry and Molecular Biology, Inc.
Figures

References
-
- Matthijs G., Schollen E., Heykants L., Grünewald S. (1999) Phosphomannomutase deficiency: the molecular basis of the classical Jaeken syndrome (CDGS type Ia). Mol. Genet. Metab. 68, 220–226 - PubMed
-
- Schollen E., Kjaergaard S., Legius E., Schwartz M., Matthijs G. (2000) Lack of Hardy-Weinberg equilibrium for the most prevalent PMM2 mutation in CDG-Ia (congenital disorders of glycosylation type Ia). Eur. J. Hum. Genet. 8, 367–371 - PubMed
-
- Kjaergaard S., Skovby F., Schwartz M. (1998) Absence of homozygosity for predominant mutations in PMM2 in Danish patients with carbohydrate-deficient glycoprotein syndrome type 1. Eur. J. Hum. Genet. 6, 331–336 - PubMed
-
- Bjursell C., Wahlström J., Berg K., Stibler H., Kristiansson B., Matthijs G., Martinsson T. (1998) Detailed mapping of the phosphomannomutase 2 (PMM2) gene and mutation detection enable improved analysis for Scandinavian CDG type I families. Eur. J. Hum. Genet. 6, 603–611 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
