

Asbestos-induced disruption of calcium homeostasis induces endoplasmic reticulum stress in macrophages
- PMID: 25324550
- PMCID: PMC4246095
- DOI: 10.1074/jbc.M114.579870
Asbestos-induced disruption of calcium homeostasis induces endoplasmic reticulum stress in macrophages
Abstract
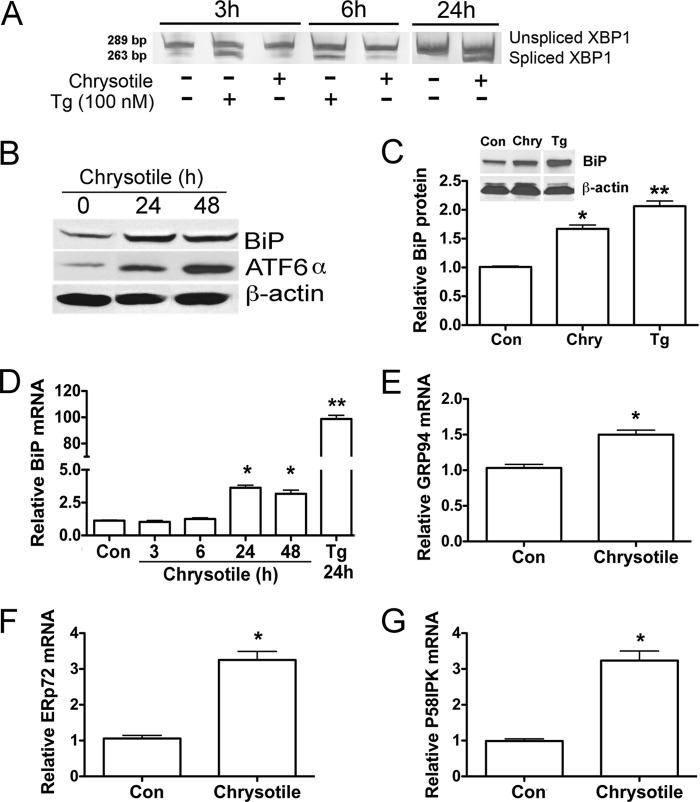
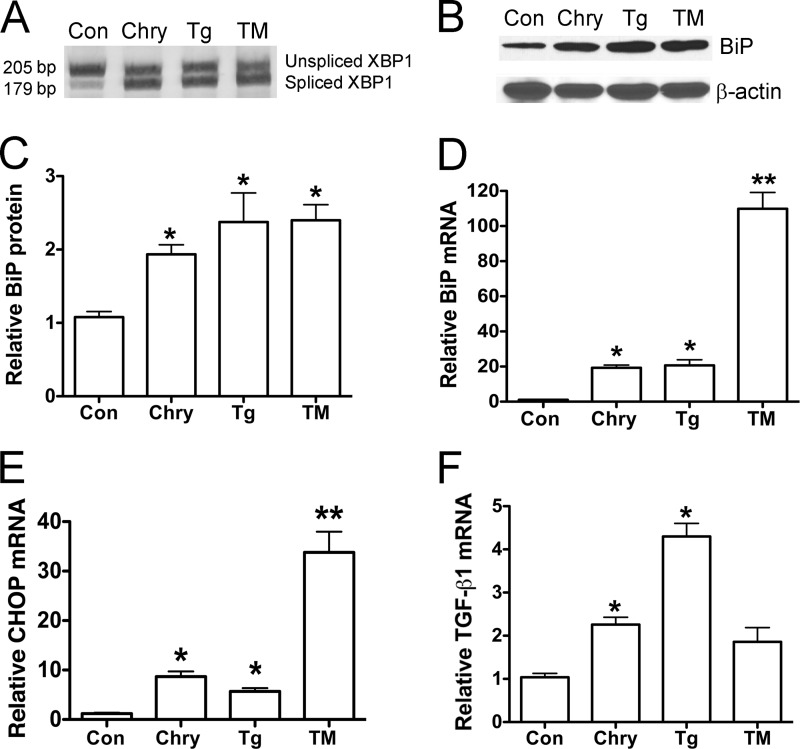
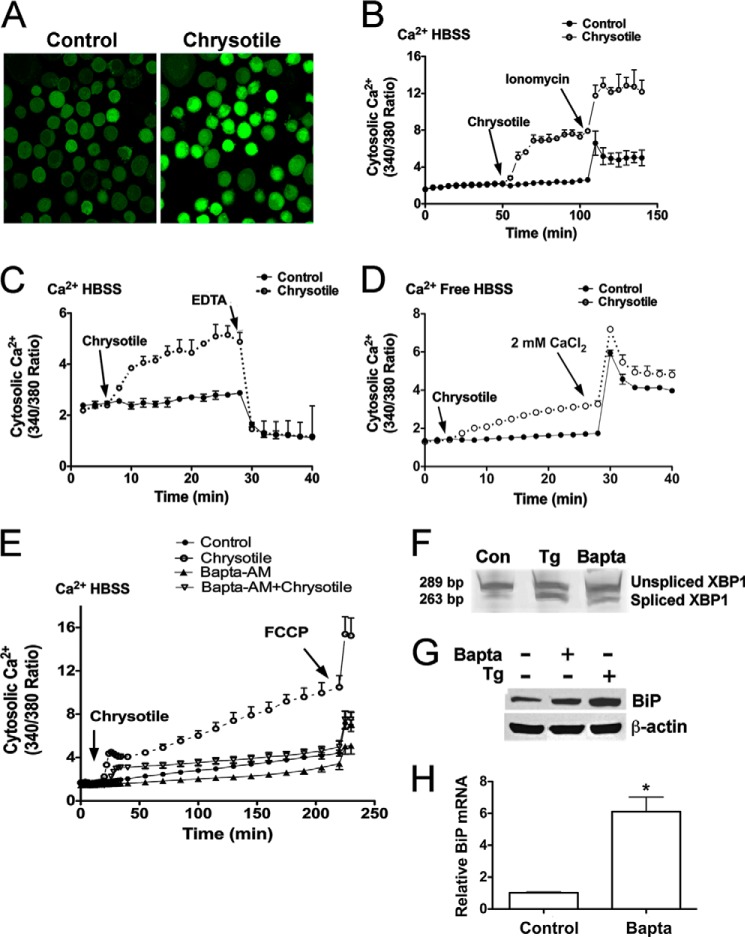
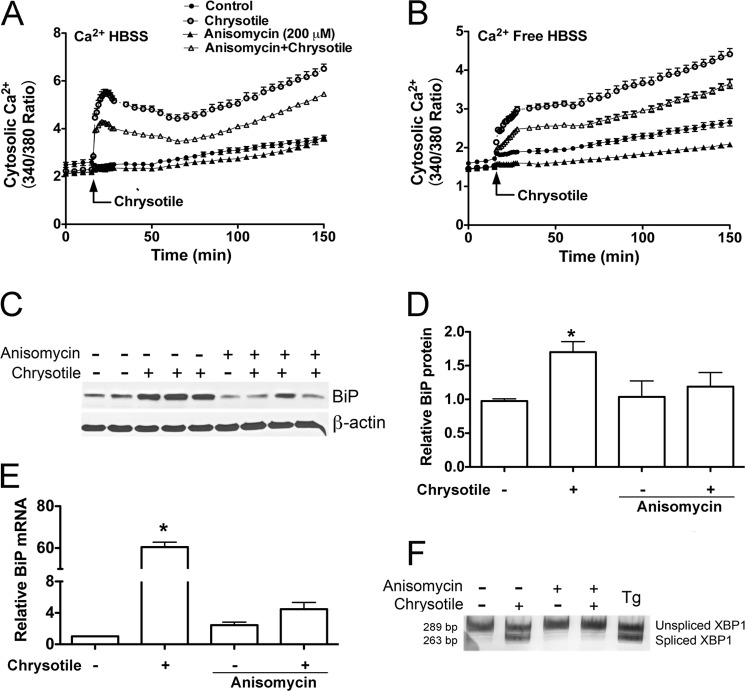
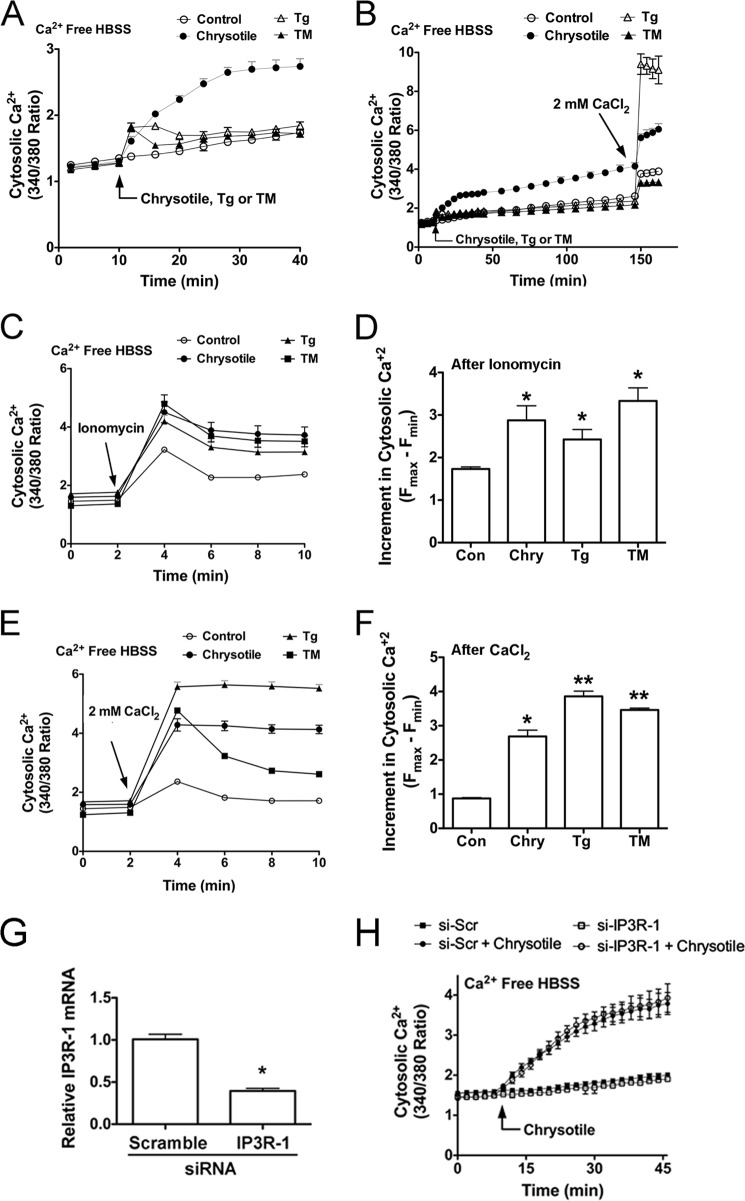
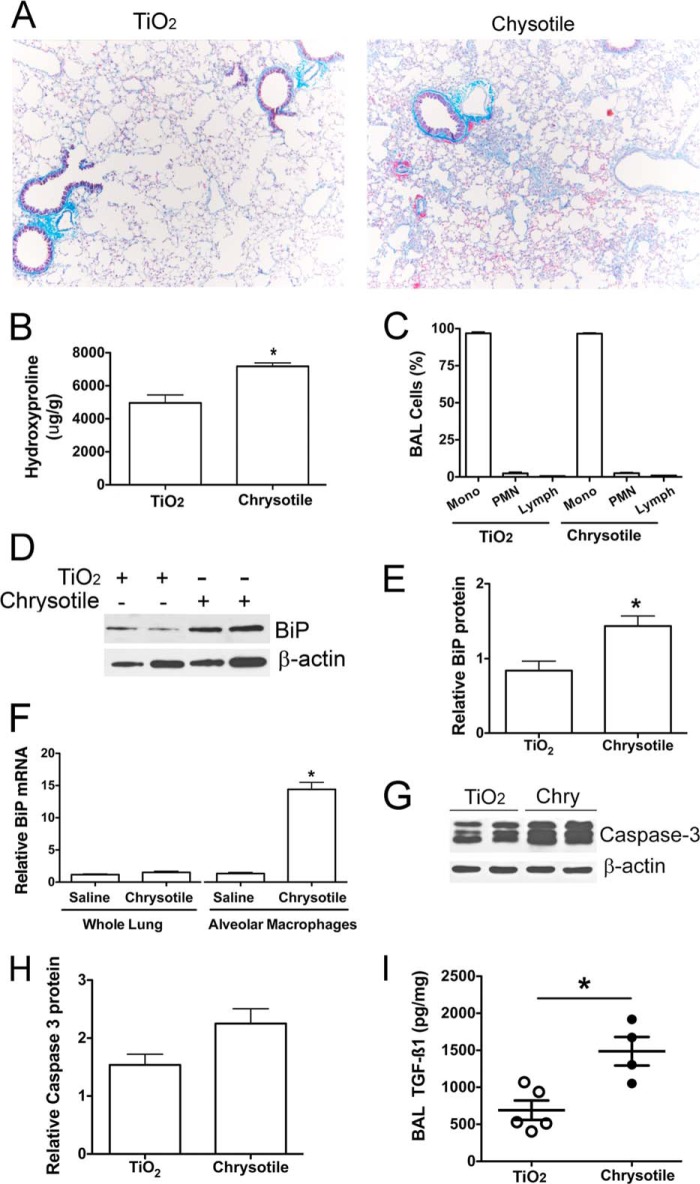
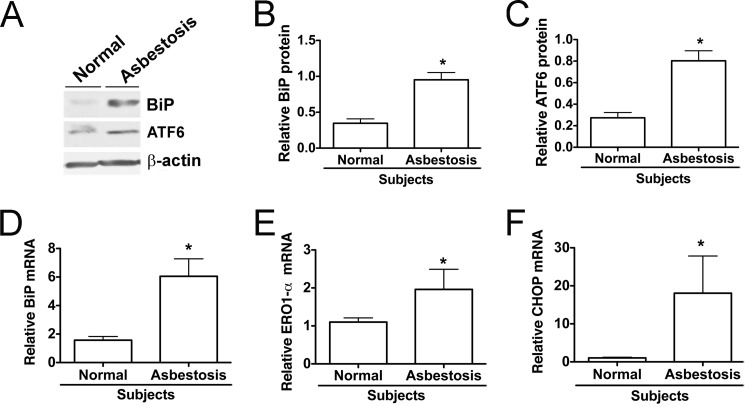
Although the mechanisms for fibrosis development remain largely unknown, recent evidence indicates that endoplasmic reticulum (ER) stress and activation of the unfolded protein response (UPR) may act as an important fibrotic stimulus in diseased lungs. ER stress is observed in lungs of patients with idiopathic pulmonary fibrosis. In this study we evaluated if ER stress and the UPR was present in macrophages exposed to chrysotile asbestos and if ER stress in macrophages was associated with asbestos-induced pulmonary fibrosis. Macrophages exposed to chrysotile had elevated transcript levels of several ER stress genes. Macrophages loaded with the Ca(2+)-sensitive dye Fura2-AM showed that cytosolic Ca(2+) increased significantly within minutes after chrysotile exposure and remained elevated for a prolonged time. Chrysotile-induced increases in cytosolic Ca(2+) were partially inhibited by either anisomycin, an inhibitor of passive Ca(2+) leak from the ER, or 1,2-bis(2-aminophenoxyl)ethane-N,N,N',N'-tetraacetic acid (BAPTA-AM), an intracellular Ca(2+) chelator known to deplete ER Ca(2+) stores. Anisomycin inhibited X-box-binding protein 1 (XBP1) mRNA splicing and reduced immunoglobulin-binding protein (BiP) levels, whereas BAPTA-AM increased XBP1 splicing and BiP expression, suggesting that ER calcium depletion may be one factor contributing to ER stress in cells exposed to chrysotile. To evaluate ER stress in vivo, asbestos-exposed mice showed fibrosis development, and alveolar macrophages from fibrotic mice showed increased expression of BiP. Bronchoalveolar macrophages from asbestosis patients showed increased expression of several ER stress genes compared with normal subjects. These findings suggest that alveolar macrophages undergo ER stress, which is associated with fibrosis development.
Keywords: Asbestos; Calcium; Endoplasmic Reticulum Stress (ER Stress); Fibrosis; Lung Injury; Macrophage; Pulmonary Fibrosis.
© 2014 by The American Society for Biochemistry and Molecular Biology, Inc.
Figures
References
-
- Guidotti T. L., Miller A., Christiani D., Wagner G., Balmes J., Harber P., Brodkin C. A., Rom W., Hillerdal G., Harbut M., Green F. H. Y. (2004) Diagnosis and initial management of nonmalignant diseases related to asbestos. Am. J. Respir. Crit. Care Med. 170, 691–715 - PubMed
-
- Hetz C. (2012) The unfolded protein response: controlling cell fate decisions under ER stress and beyond. Nat. Rev. Mol. Cell Biol. 13, 89–102 - PubMed
-
- Kim I., Xu W., Reed J. C. (2008) Cell death and endoplasmic reticulum stress: disease relevance and therapeutic opportunities. Nat. Rev. Drug Discov. 7, 1013–1030 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
Miscellaneous
