

Speciation of iron in mouse liver during development, iron deficiency, IRP2 deletion and inflammatory hepatitis
- PMID: 25325718
- PMCID: PMC4276432
- DOI: 10.1039/c4mt00215f
Speciation of iron in mouse liver during development, iron deficiency, IRP2 deletion and inflammatory hepatitis
Abstract
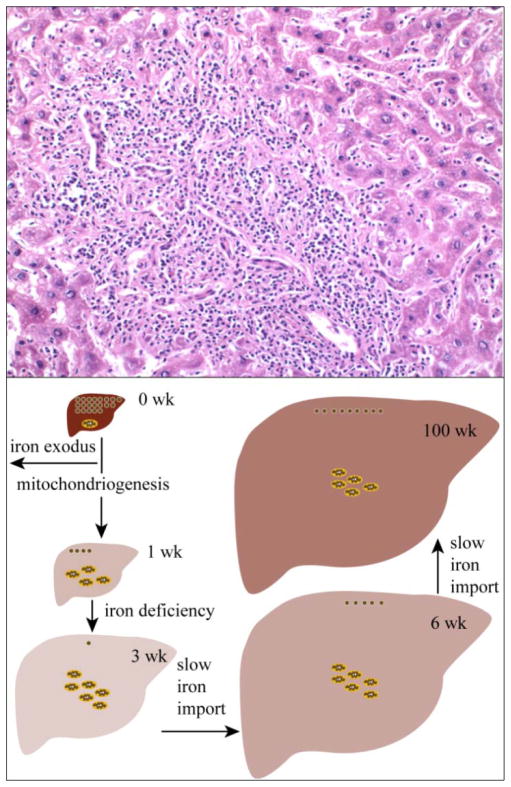
The iron content of livers from (57)Fe-enriched C57BL/6 mice of different ages were investigated using Mössbauer spectroscopy, electron paramagnetic resonance (EPR), electronic absorption spectroscopy and inductively coupled plasma mass spectrometry (ICP-MS). About 80% of the Fe in an adult liver was due to blood; thus removal of blood by flushing with buffer was essential to observe endogenous liver Fe. Even after exhaustive flushing, ca. 20% of the Fe in anaerobically dissected livers was typical of deoxy-hemoglobin. The concentration of Fe in newborn livers was the highest of any developmental stage (∼1.2 mM). Most was stored as ferritin, with little mitochondrial Fe (consisting primarily of Fe-S clusters and haems) evident. Within the first few weeks of life, about half of ferritin Fe was mobilized and exported, illustrating the importance of Fe release as well as Fe storage in liver function. Additional ferritin Fe was used to generate mitochondrial Fe centres. From ca. 4 weeks of age to the end of the mouse's natural lifespan, the concentration of mitochondrial Fe in liver was essentially invariant. A minor contribution from nonhaem high-spin Fe(II) was observed in most liver samples and was also invariant with age. Some portion of these species may constitute the labile iron pool. Livers from mice raised on an Fe-deficient diet were highly Fe depleted; they were devoid of ferritin and contained 1/3 as much mitochondrial Fe as found in Fe-sufficient livers. In contrast, brains of the same Fe-deficient mice retained normal levels of mitochondrial Fe. Livers from mice with inflammatory hepatitis and from IRP2(-/-) mice hyper-accumulated Fe. These livers had high ferritin levels but low levels of mitochondrial Fe.
Figures
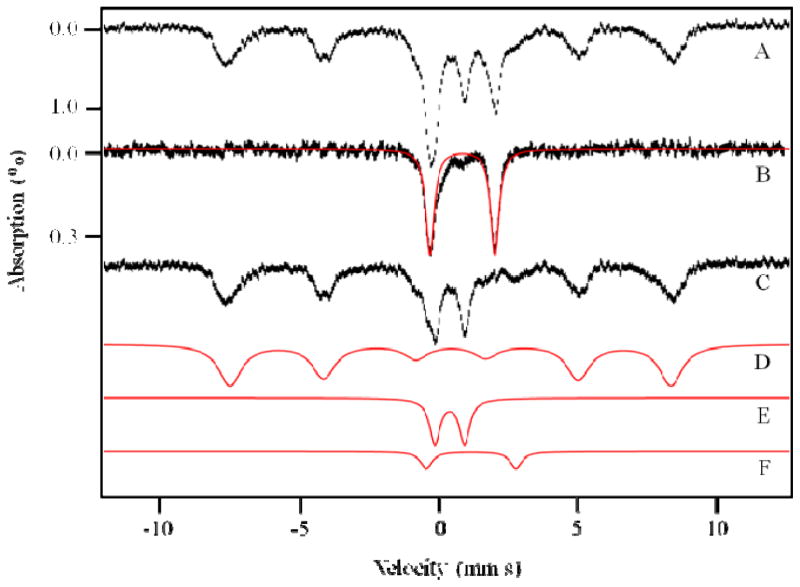
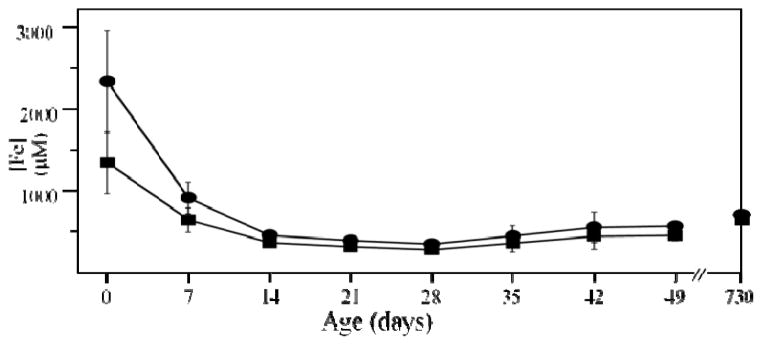
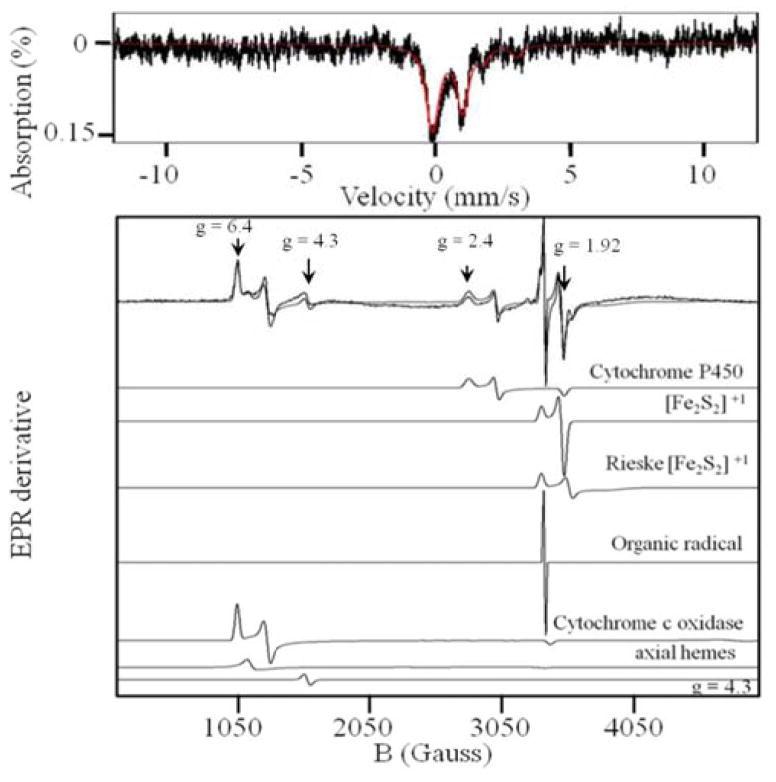
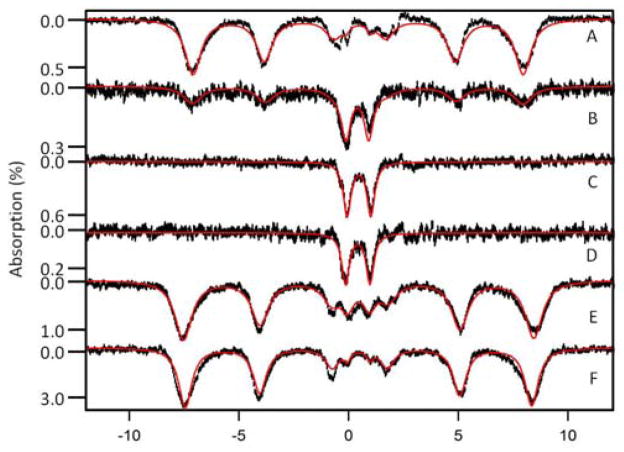
References
-
- Ganz T. Systemic Iron Homeostasis. Physiol Rev. 2013;93:1721–1741. - PubMed
-
- Anderson GJ, Frazer DM. Hepatic Iron Metabolism. Seminars in Liver Disease. 2005;25:420–432. - PubMed
-
- Ganz T, Vaulont S. Molecular Regulation of Systemic Iron Metabolism. In: Anderson GJ, McLaren GD, editors. Iron Physiology and Pathophysiology in Humans. Humana Press; NJ: 2012. pp. 173–190.
-
- Andrews SC. The Ferritin-like superfamily: evolution of the biological iron storeman from a rubrerythrin-like ancestor. Biochem Biophys Acta General Subjects. 2010;1800:691–705. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous
