

Overelaborated synaptic architecture and reduced synaptomatrix glycosylation in a Drosophila classic galactosemia disease model
- PMID: 25326312
- PMCID: PMC4257005
- DOI: 10.1242/dmm.017137
Overelaborated synaptic architecture and reduced synaptomatrix glycosylation in a Drosophila classic galactosemia disease model
Abstract
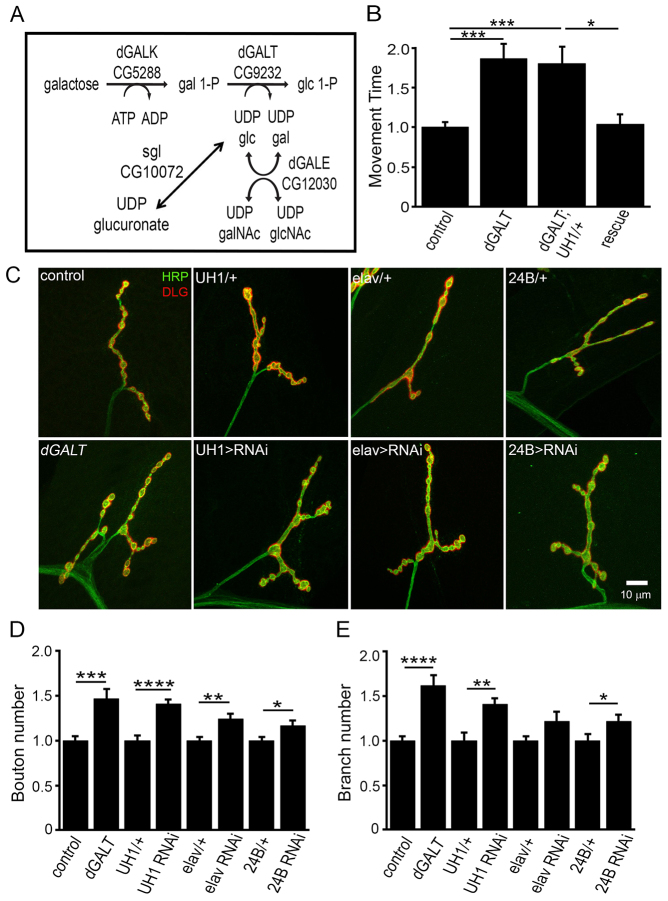
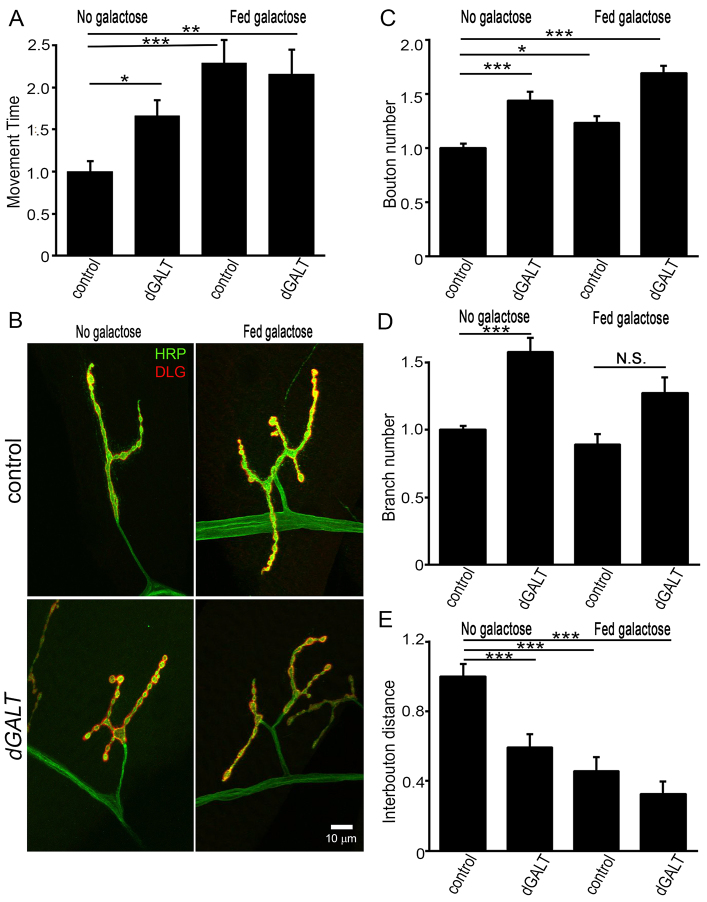
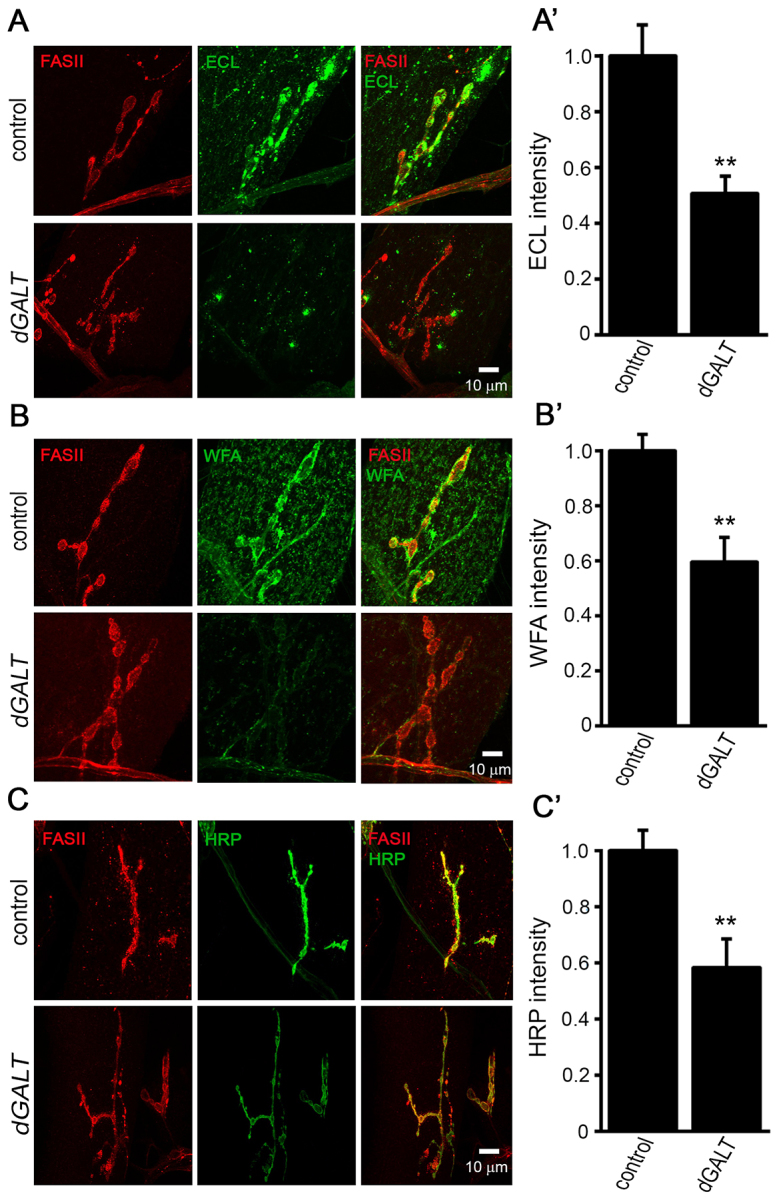
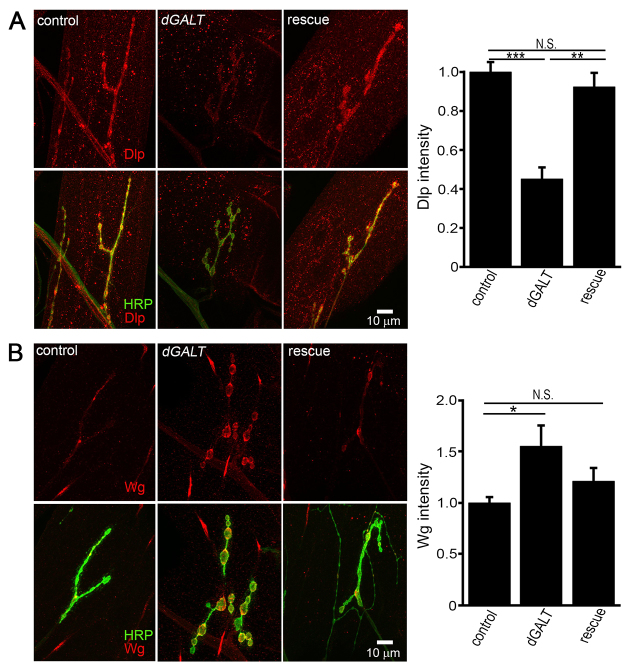
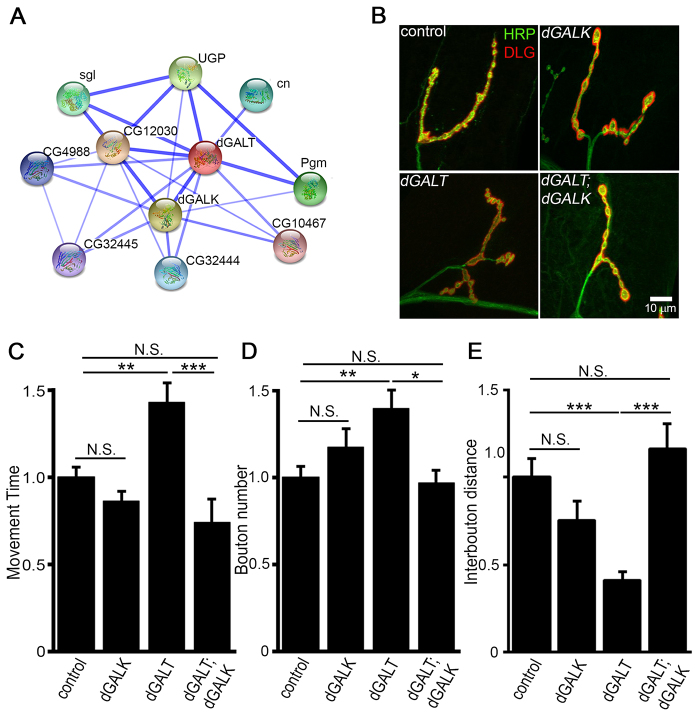
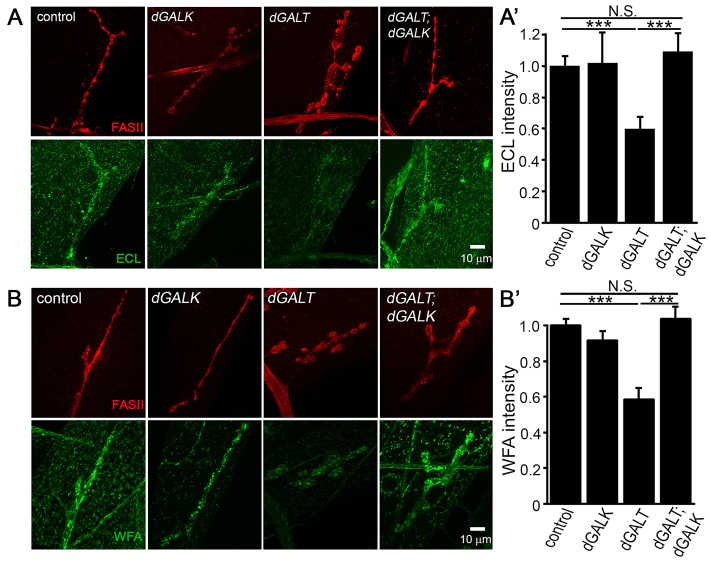
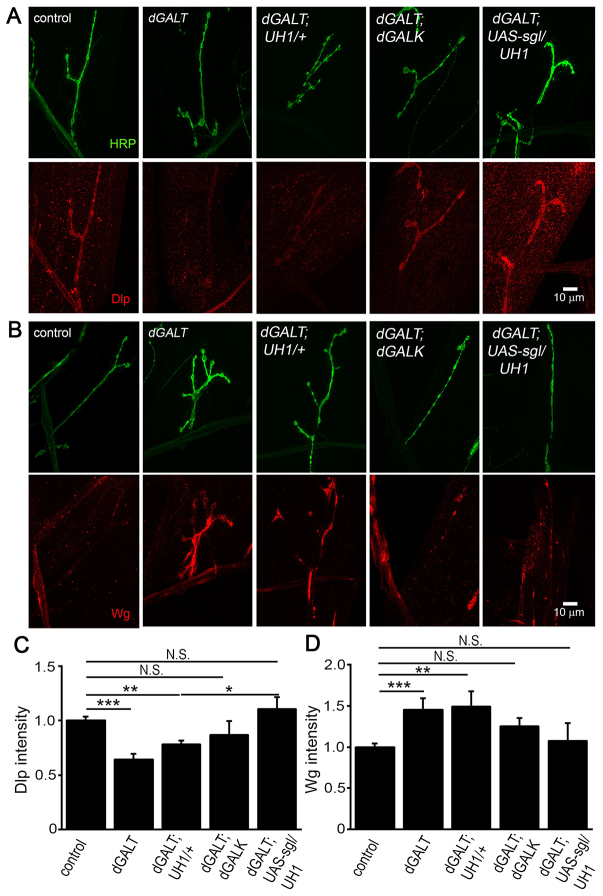
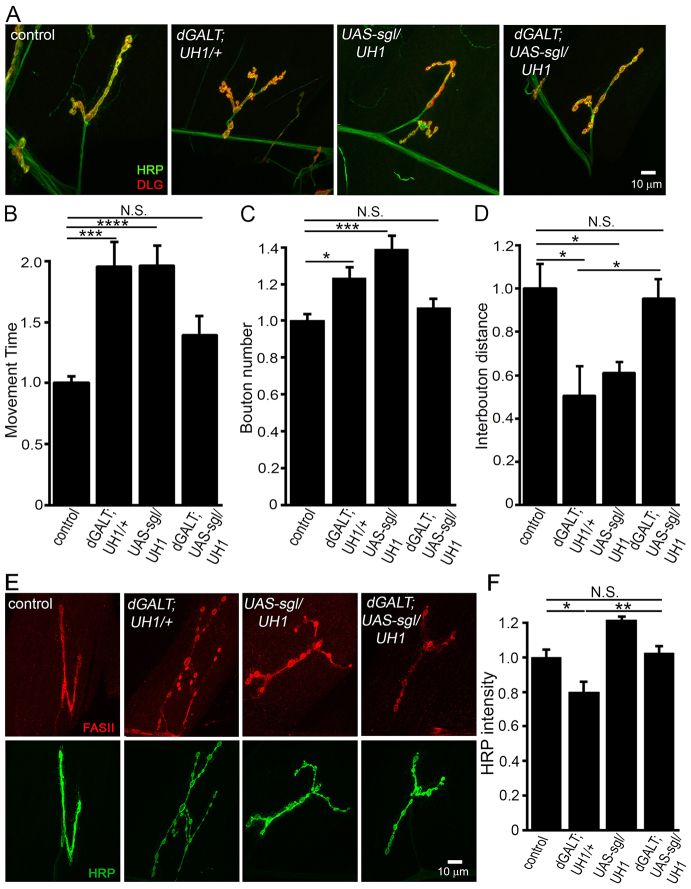
Classic galactosemia (CG) is an autosomal recessive disorder resulting from loss of galactose-1-phosphate uridyltransferase (GALT), which catalyzes conversion of galactose-1-phosphate and uridine diphosphate (UDP)-glucose to glucose-1-phosphate and UDP-galactose, immediately upstream of UDP-N-acetylgalactosamine and UDP-N-acetylglucosamine synthesis. These four UDP-sugars are essential donors for driving the synthesis of glycoproteins and glycolipids, which heavily decorate cell surfaces and extracellular spaces. In addition to acute, potentially lethal neonatal symptoms, maturing individuals with CG develop striking neurodevelopmental, motor and cognitive impairments. Previous studies suggest that neurological symptoms are associated with glycosylation defects, with CG recently being described as a congenital disorder of glycosylation (CDG), showing defects in both N- and O-linked glycans. Here, we characterize behavioral traits, synaptic development and glycosylated synaptomatrix formation in a GALT-deficient Drosophila disease model. Loss of Drosophila GALT (dGALT) greatly impairs coordinated movement and results in structural overelaboration and architectural abnormalities at the neuromuscular junction (NMJ). Dietary galactose and mutation of galactokinase (dGALK) or UDP-glucose dehydrogenase (sugarless) genes are identified, respectively, as critical environmental and genetic modifiers of behavioral and cellular defects. Assaying the NMJ extracellular synaptomatrix with a broad panel of lectin probes reveals profound alterations in dGALT mutants, including depletion of galactosyl, N-acetylgalactosamine and fucosylated horseradish peroxidase (HRP) moieties, which are differentially corrected by dGALK co-removal and sugarless overexpression. Synaptogenesis relies on trans-synaptic signals modulated by this synaptomatrix carbohydrate environment, and dGALT-null NMJs display striking changes in heparan sulfate proteoglycan (HSPG) co-receptor and Wnt ligand levels, which are also corrected by dGALK co-removal and sugarless overexpression. These results reveal synaptomatrix glycosylation losses, altered trans-synaptic signaling pathway components, defective synaptogenesis and impaired coordinated movement in a CG neurological disease model.
Keywords: Congenital disorder of glycosylation (CDG); Galactokinase; HSPG; Neuromuscular junction; Synaptogenesis; Trans-synaptic signaling; WNT; sugarless.
© 2014. Published by The Company of Biologists Ltd.
Figures
Similar articles
-
Coordinated movement, neuromuscular synaptogenesis and trans-synaptic signaling defects in Drosophila galactosemia models.Hum Mol Genet. 2016 Sep 1;25(17):3699-3714. doi: 10.1093/hmg/ddw217. Epub 2016 Jul 27. Hum Mol Genet. 2016. PMID: 27466186 Free PMC article.
-
Synaptic roles for phosphomannomutase type 2 in a new Drosophila congenital disorder of glycosylation disease model.Dis Model Mech. 2016 May 1;9(5):513-27. doi: 10.1242/dmm.022939. Epub 2016 Mar 3. Dis Model Mech. 2016. PMID: 26940433 Free PMC article.
-
N-glycosylation requirements in neuromuscular synaptogenesis.Development. 2013 Dec;140(24):4970-81. doi: 10.1242/dev.099192. Epub 2013 Nov 13. Development. 2013. PMID: 24227656 Free PMC article.
-
Galactosemia: when is it a newborn screening emergency?Mol Genet Metab. 2012 May;106(1):7-11. doi: 10.1016/j.ymgme.2012.03.007. Epub 2012 Mar 21. Mol Genet Metab. 2012. PMID: 22483615 Review.
-
Glycosylated synaptomatrix regulation of trans-synaptic signaling.Dev Neurobiol. 2012 Jan;72(1):2-21. doi: 10.1002/dneu.20891. Dev Neurobiol. 2012. PMID: 21509945 Free PMC article. Review.
Cited by
-
Fragment Screening Reveals Starting Points for Rational Design of Galactokinase 1 Inhibitors to Treat Classic Galactosemia.ACS Chem Biol. 2021 Apr 16;16(4):586-595. doi: 10.1021/acschembio.0c00498. Epub 2021 Mar 16. ACS Chem Biol. 2021. PMID: 33724769 Free PMC article.
-
Neuron-to-glia and glia-to-glia signaling directs critical period experience-dependent synapse pruning.Front Cell Dev Biol. 2025 Feb 18;13:1540052. doi: 10.3389/fcell.2025.1540052. eCollection 2025. Front Cell Dev Biol. 2025. PMID: 40040788 Free PMC article. Review.
-
Sweet and sour: an update on classic galactosemia.J Inherit Metab Dis. 2017 May;40(3):325-342. doi: 10.1007/s10545-017-0029-3. Epub 2017 Mar 9. J Inherit Metab Dis. 2017. PMID: 28281081 Free PMC article. Review.
-
Extracellular heparan sulfate proteoglycans and glycan-binding lectins orchestrate trans-synaptic signaling.J Cell Sci. 2020 Aug 11;133(15):jcs244186. doi: 10.1242/jcs.244186. J Cell Sci. 2020. PMID: 32788209 Free PMC article. Review.
-
Classical Galactosaemia and CDG, the N-Glycosylation Interface. A Review.JIMD Rep. 2017;34:33-42. doi: 10.1007/8904_2016_5. Epub 2016 Aug 9. JIMD Rep. 2017. PMID: 27502837 Free PMC article.
References
-
- Andlauer T. F., Sigrist S. J. (2012). In vivo imaging of the Drosophila larval neuromuscular junction. Cold Spring Harb. Protoc. 2012, 481–489. - PubMed
-
- Binari R. C., Staveley B. E., Johnson W. A., Godavarti R., Sasisekharan R., Manoukian A. S. (1997). Genetic evidence that heparin-like glycosaminoglycans are involved in wingless signaling. Development 124, 2623–2632. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
