

Nucleotide synthesis is regulated by cytoophidium formation during neurodevelopment and adaptive metabolism
- PMID: 25326513
- PMCID: PMC4232762
- DOI: 10.1242/bio.201410165
Nucleotide synthesis is regulated by cytoophidium formation during neurodevelopment and adaptive metabolism
Abstract
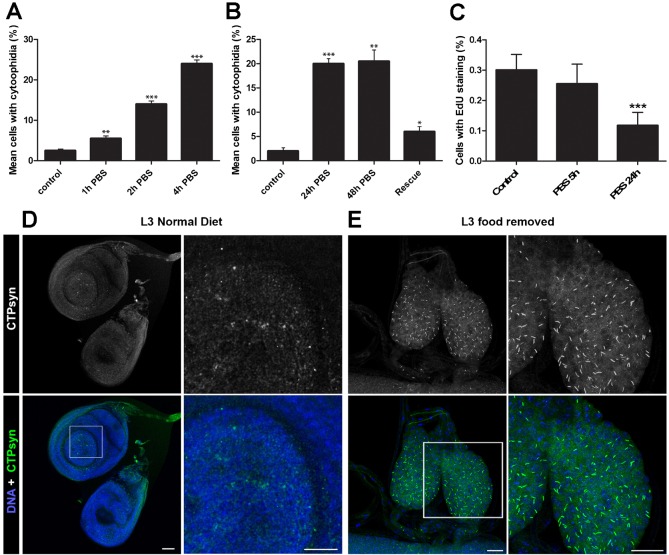
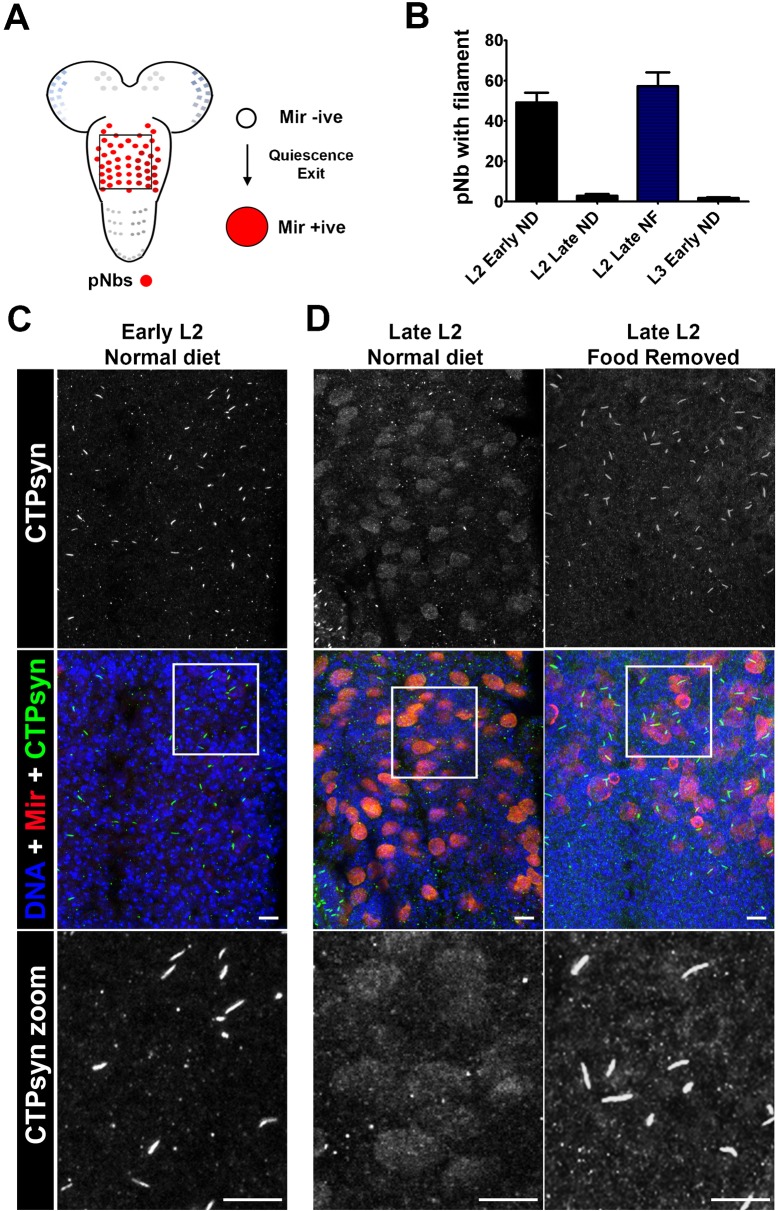
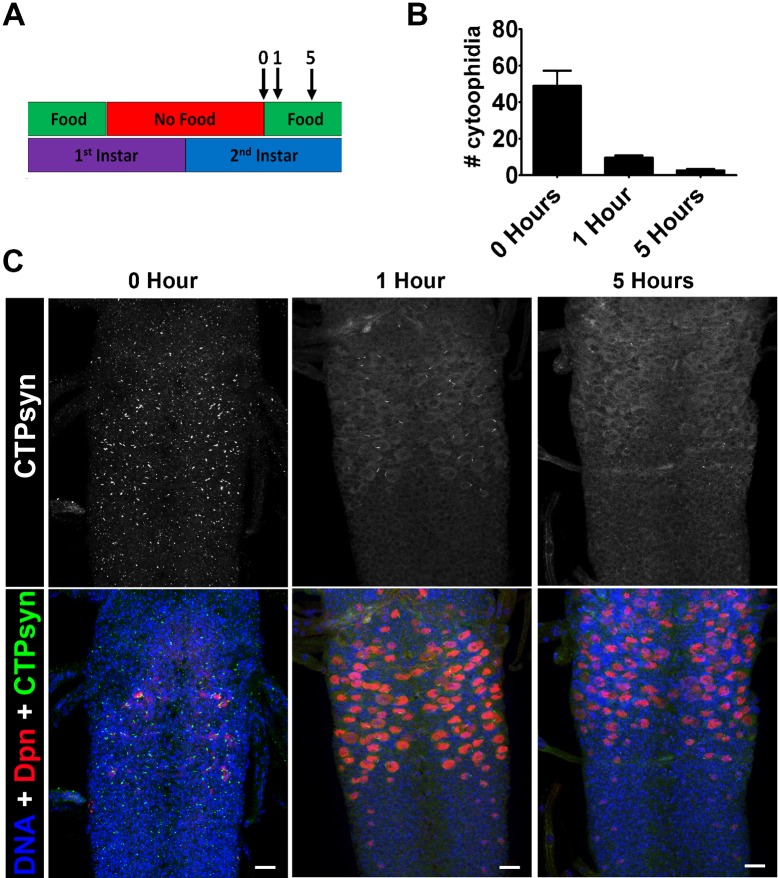
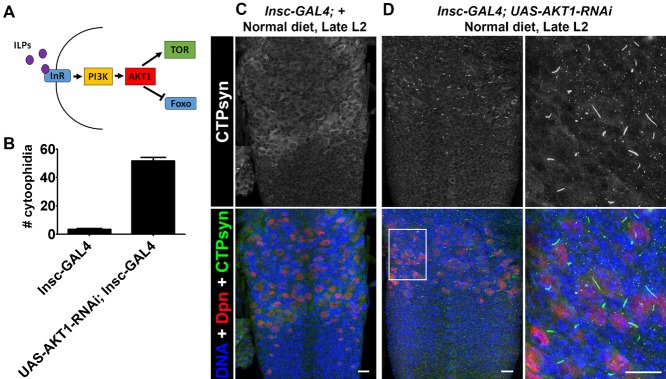
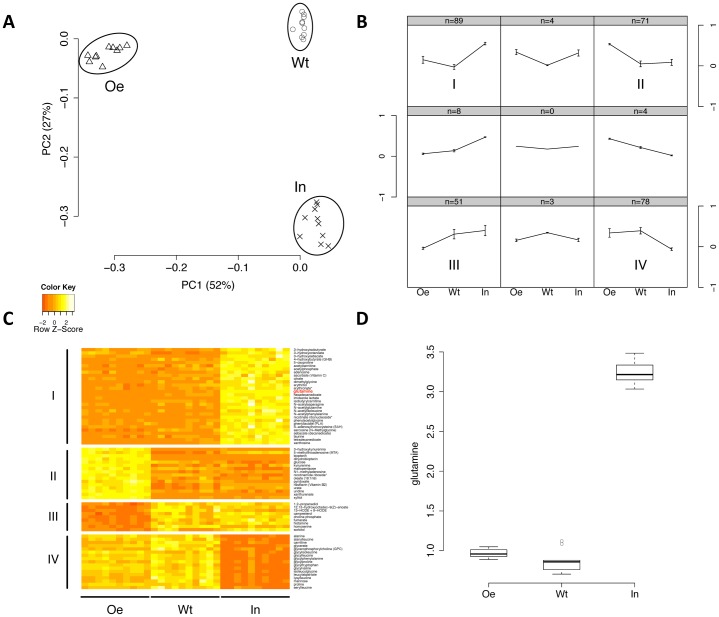
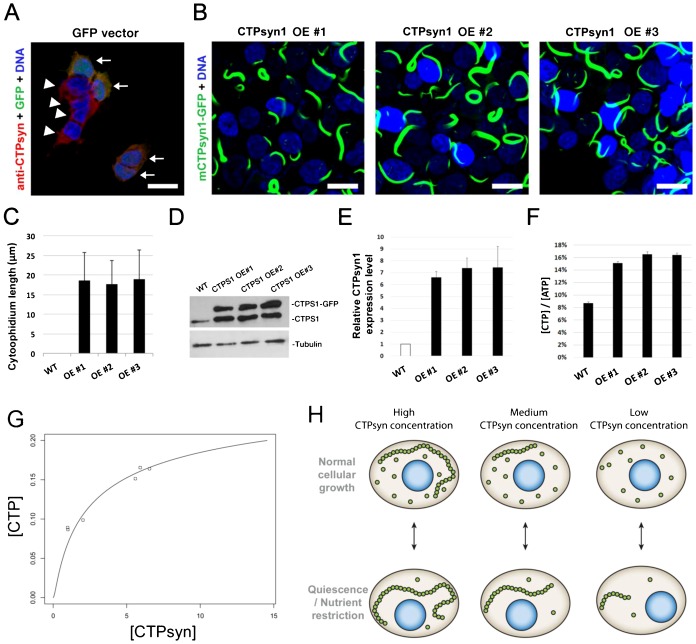
The essential metabolic enzyme CTP synthase (CTPsyn) can be compartmentalised to form an evolutionarily-conserved intracellular structure termed the cytoophidium. Recently, it has been demonstrated that the enzymatic activity of CTPsyn is attenuated by incorporation into cytoophidia in bacteria and yeast cells. Here we demonstrate that CTPsyn is regulated in a similar manner in Drosophila tissues in vivo. We show that cytoophidium formation occurs during nutrient deprivation in cultured cells, as well as in quiescent and starved neuroblasts of the Drosophila larval central nervous system. We also show that cytoophidia formation is reversible during neurogenesis, indicating that filament formation regulates pyrimidine synthesis in a normal developmental context. Furthermore, our global metabolic profiling demonstrates that CTPsyn overexpression does not significantly alter CTPsyn-related enzymatic activity, suggesting that cytoophidium formation facilitates metabolic stabilisation. In addition, we show that overexpression of CTPsyn only results in moderate increase of CTP pool in human stable cell lines. Together, our study provides experimental evidence, and a mathematical model, for the hypothesis that inactive CTPsyn is incorporated into cytoophidia.
Keywords: CTP; CTP synthase; Drosophila; cytoophidium; intracellular compartmentation; neurogenesis.
© 2014. Published by The Company of Biologists Ltd.
Conflict of interest statement
Figures
References
-
- Aronow B., Ullman B. (1987). In situ regulation of mammalian CTP synthetase by allosteric inhibition. J. Biol. Chem. 262, 5106–5112. - PubMed
-
- Beaty N. B., Lane M. D. (1983a). Kinetics of activation of acetyl-CoA carboxylase by citrate. Relationship to the rate of polymerization of the enzyme. J. Biol. Chem. 258, 13043–13050. - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
