

Nectin family of cell-adhesion molecules: structural and molecular aspects of function and specificity
- PMID: 25326769
- PMCID: PMC11113404
- DOI: 10.1007/s00018-014-1763-4
Nectin family of cell-adhesion molecules: structural and molecular aspects of function and specificity
Abstract
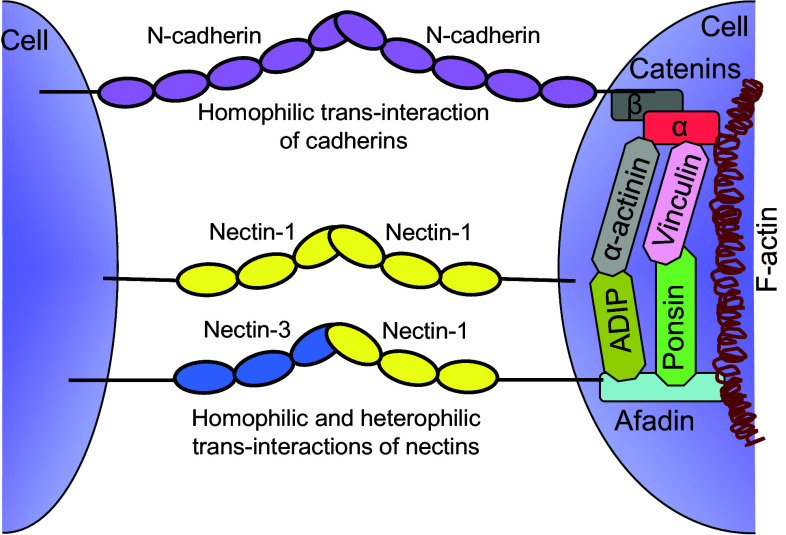
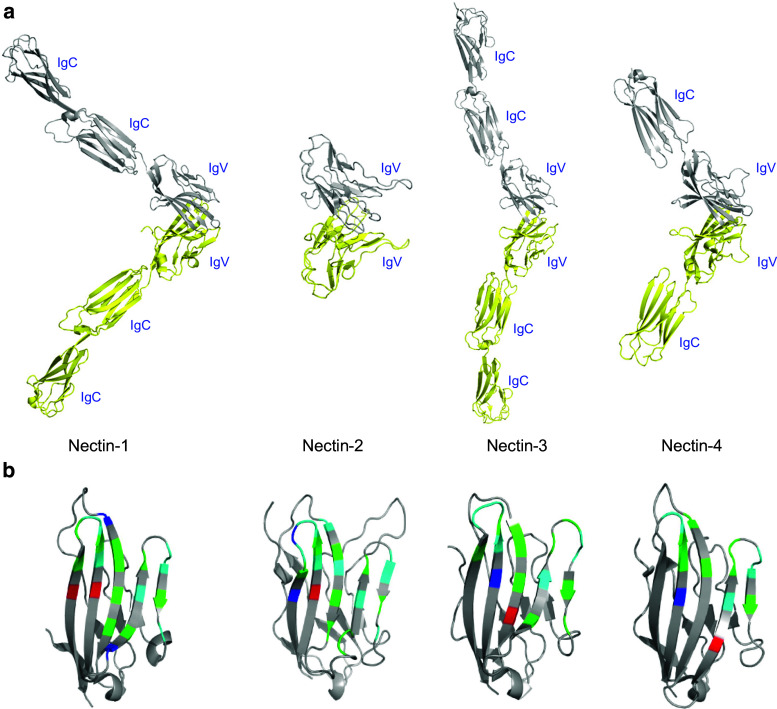
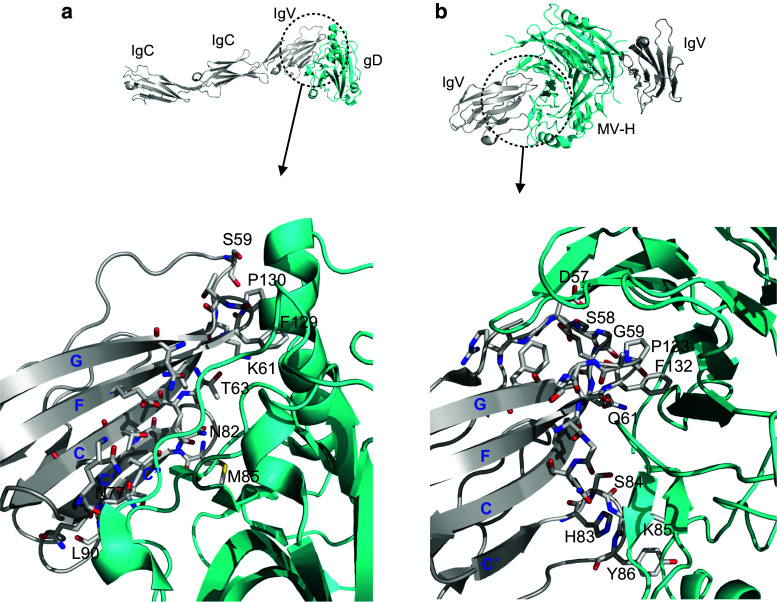
Cell-cell adhesive processes are central to the physiology of multicellular organisms. A number of cell surface molecules contribute to cell-cell adhesion, and the dysfunction of adhesive processes underlies numerous developmental defects and inherited diseases. The nectins, a family of four immunoglobulin superfamily members (nectin-1 to -4), interact through their extracellular domains to support cell-cell adhesion. While both homophilic and heterophilic interactions among the nectins are implicated in cell-cell adhesion, cell-based and biochemical studies suggest heterophilic interactions are stronger than homophilic interactions and control a range of physiological processes. In addition to interactions within the nectin family, heterophilic associations with nectin-like molecules, immune receptors, and viral glycoproteins support a wide range of biological functions, including immune modulation, cancer progression, host-pathogen interactions and immune evasion. We review current structural and molecular knowledge of nectin recognition processes, with a focus on the biochemical and biophysical determinants of affinity and selectivity that drive distinct nectin associations. These proteins and interactions are discussed as potential targets for immunotherapy.
Figures





References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
