

Regulation of PKC mediated signaling by calcium during visceral leishmaniasis
- PMID: 25329062
- PMCID: PMC4201563
- DOI: 10.1371/journal.pone.0110843
Regulation of PKC mediated signaling by calcium during visceral leishmaniasis
Abstract
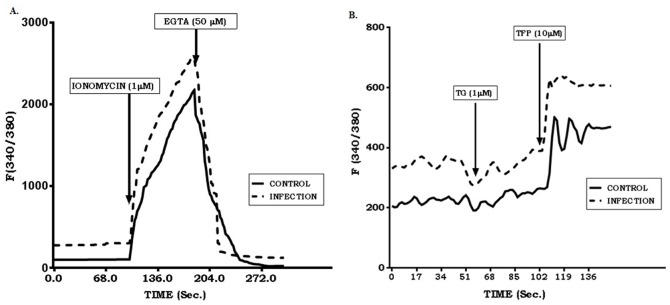
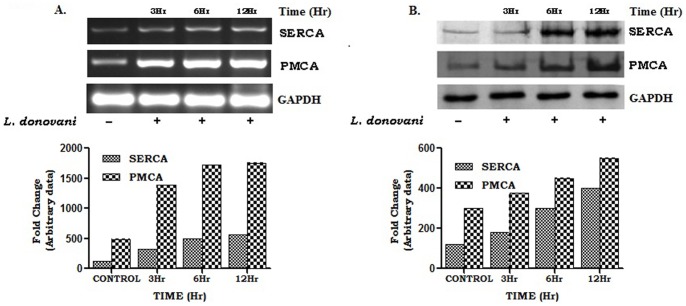
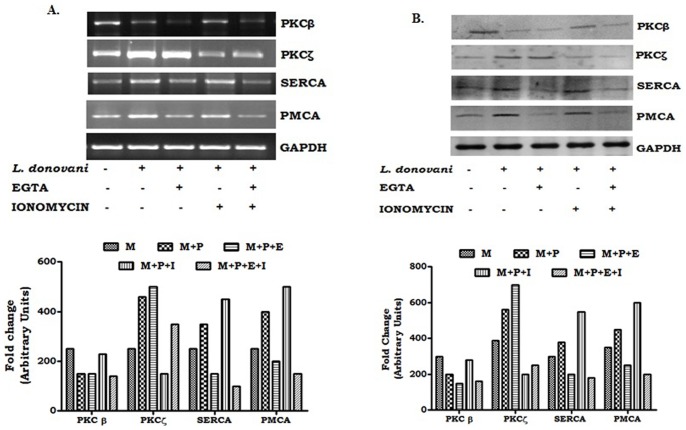
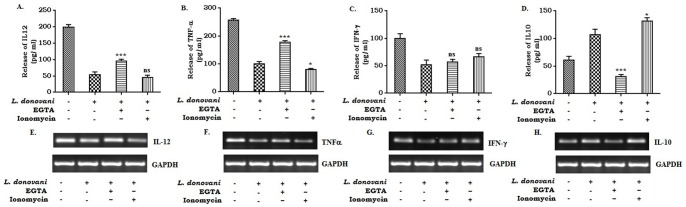
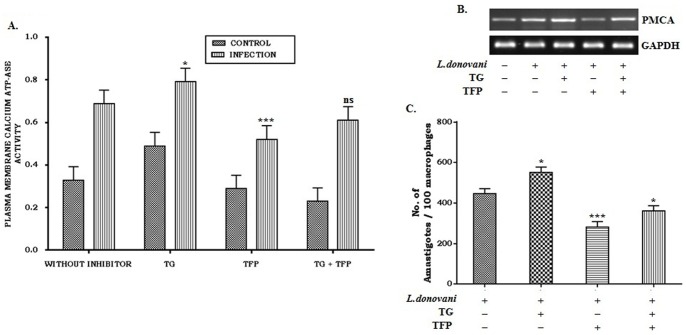
Calcium is an ubiquitous cellular signaling molecule that controls a variety of cellular processes and is strictly maintained in the cellular compartments by the coordination of various Ca2+ pumps and channels. Two such fundamental calcium pumps are plasma membrane calcium ATPase (PMCA) and Sarco/endoplasmic reticulum calcium ATPase (SERCA) which play a pivotal role in maintaining intracellular calcium homeostasis. This intracellular Ca2+ homeostasis is often disturbed by the protozoan parasite Leishmania donovani, the causative organism of visceral leishmaniasis. In the present study we have dileneated the involvement of PMCA4 and SERCA3 during leishmaniasis. We have observed that during leishmaniasis, intracellular Ca2+ concentration was up-regulated and was further controlled by both PMCA4 and SERCA3. Inhibition of these two Ca2+-ATPases resulted in decreased parasite burden within the host macrophages due to enhanced intracellular Ca2+. Contrastingly, on the other hand, activation of PMCA4 was found to enhance the parasite burden. Our findings also highlighted the importance of Ca2+ in the modulation of cytokine balance during leishmaniasis. These results thus cumulatively suggests that these two Ca2+-ATPases play prominent roles during visceral leishmaniasis.
Conflict of interest statement
Figures
References
-
- Murray HW, Spitalny GL, Nathan CF (1985) Activation of mouse peritoneal macrophages in vitro and in vivo by interferon-gamma. J Immunol 134: 1619–1622. - PubMed
-
- Wilmann M, Gautel M, Mayans O (2000) Activation of calcium/calmodulin regulated kinases. Cell Mol Biol 46(5): 883–94. - PubMed
-
- Nishizuka (1988) Studies and prospectives of protein kinase C in signal transduction. Nippon Ketsueki Gakkai Zasshi 51(8): 1321–6. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
