

Lipid kinases are essential for apicoplast homeostasis in Toxoplasma gondii
- PMID: 25329540
- PMCID: PMC4356647
- DOI: 10.1111/cmi.12383
Lipid kinases are essential for apicoplast homeostasis in Toxoplasma gondii
Abstract
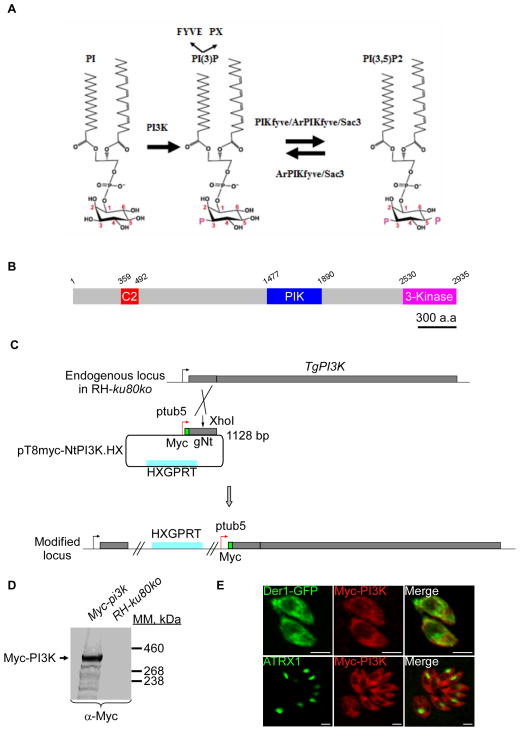
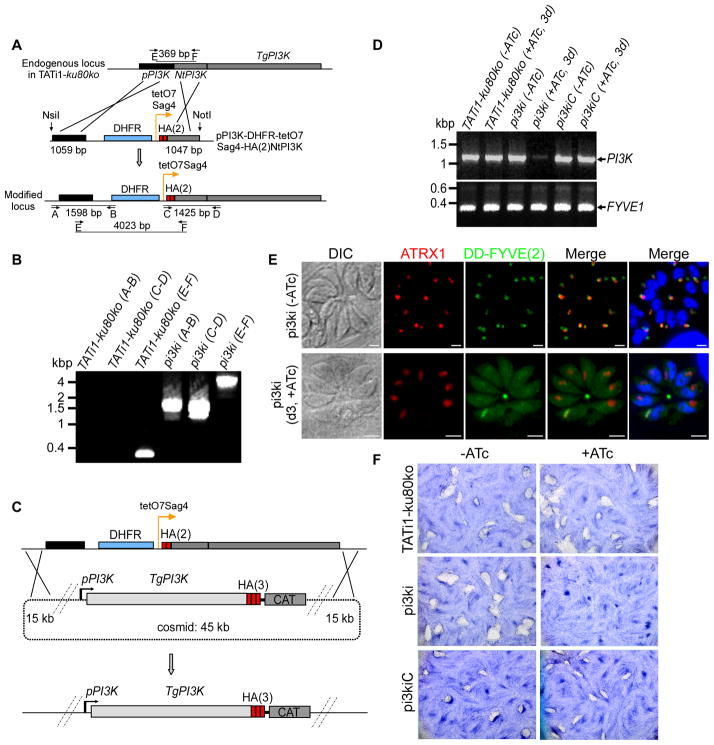
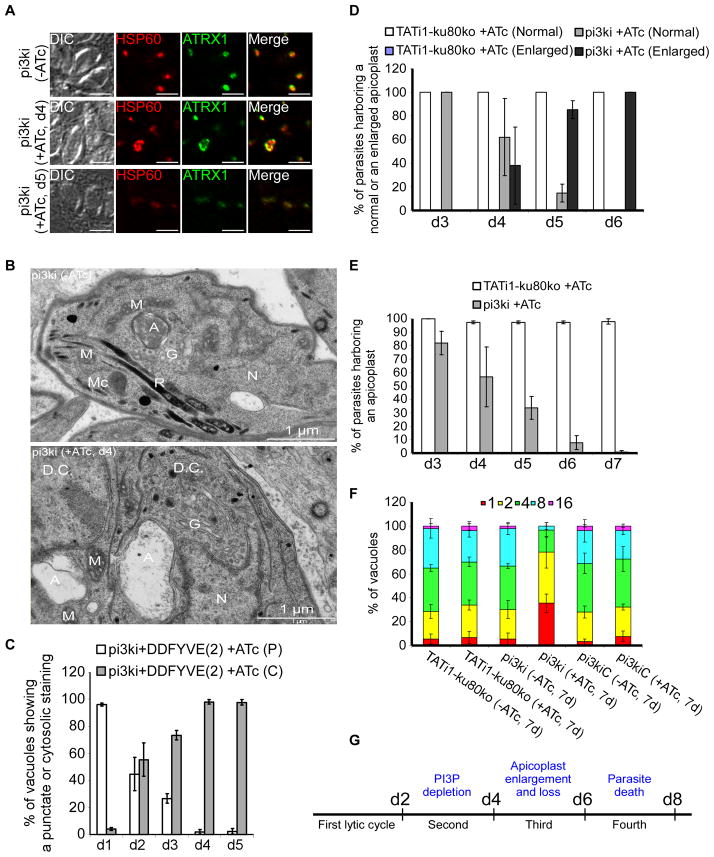
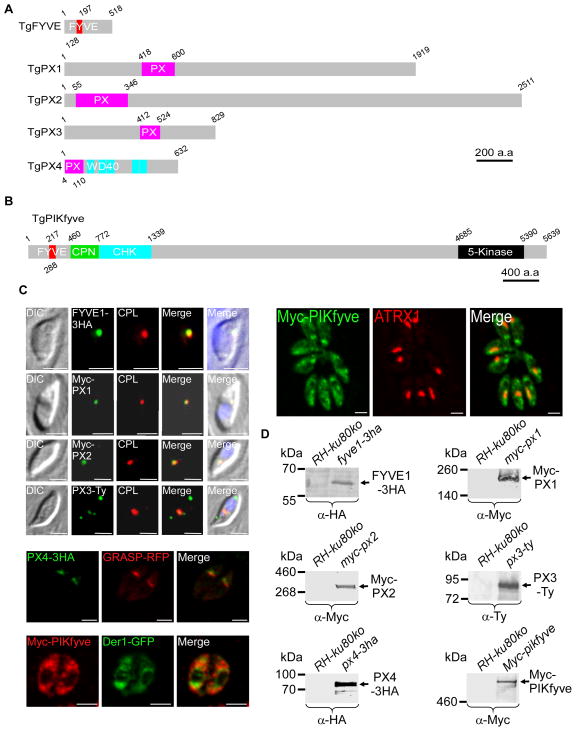
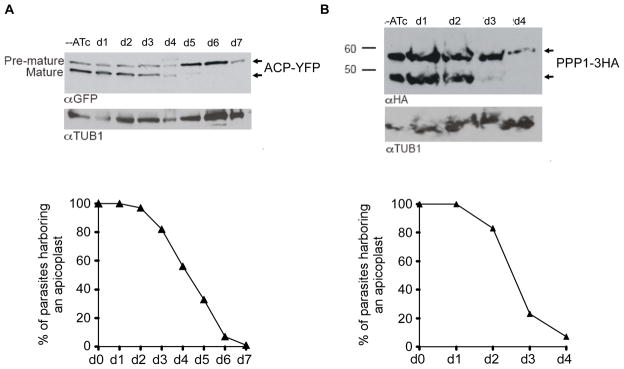
Phosphoinositides regulate numerous cellular processes by recruiting cytosolic effector proteins and acting as membrane signalling entities. The cellular metabolism and localization of phosphoinositides are tightly regulated by distinct lipid kinases and phosphatases. Here, we identify and characterize a unique phosphatidylinositol 3 kinase (PI3K) in Toxoplasma gondii, a protozoan parasite belonging to the phylum Apicomplexa. Conditional depletion of this enzyme and subsequently of its product, PI(3)P, drastically alters the morphology and inheritance of the apicoplast, an endosymbiotic organelle of algal origin that is a unique feature of many Apicomplexa. We searched the T. gondii genome for PI(3)P-binding proteins and identified in total six PX and FYVE domain-containing proteins including a PIKfyve lipid kinase, which phosphorylates PI(3)P into PI(3,5)P2 . Although depletion of putative PI(3)P-binding proteins shows that they are not essential for parasite growth and apicoplast biology, conditional disruption of PIKfyve induces enlarged apicoplasts, as observed upon loss of PI(3)P. A similar defect of apicoplast homeostasis was also observed by knocking down the PIKfyve regulatory protein ArPIKfyve, suggesting that in T. gondii, PI(3)P-related function for the apicoplast might mainly be to serve as a precursor for the synthesis of PI(3,5)P2 . Accordingly, PI3K is conserved in all apicomplexan parasites whereas PIKfyve and ArPIKfyve are absent in Cryptosporidium species that lack an apicoplast, supporting a direct role of PI(3,5)P2 in apicoplast homeostasis. This study enriches the already diverse functions attributed to PI(3,5)P2 in eukaryotic cells and highlights these parasite lipid kinases as potential drug targets.
© 2014 John Wiley & Sons Ltd.
Figures
Similar articles
-
Autophagy-Related Protein ATG18 Regulates Apicoplast Biogenesis in Apicomplexan Parasites.mBio. 2017 Oct 31;8(5):e01468-17. doi: 10.1128/mBio.01468-17. mBio. 2017. PMID: 29089429 Free PMC article.
-
Toxoplasma TgAtg8-TgAtg3 Interaction Primarily Contributes to Apicoplast Inheritance and Parasite Growth in Tachyzoite.Microbiol Spectr. 2022 Feb 23;10(1):e0149521. doi: 10.1128/spectrum.01495-21. Epub 2022 Feb 23. Microbiol Spectr. 2022. PMID: 35196797 Free PMC article.
-
Characterization of the apicoplast-localized enzyme TgUroD in Toxoplasma gondii reveals a key role of the apicoplast in heme biosynthesis.J Biol Chem. 2020 Feb 7;295(6):1539-1550. doi: 10.1074/jbc.RA119.011605. Epub 2019 Dec 30. J Biol Chem. 2020. PMID: 31914409 Free PMC article.
-
The metabolic pathways and transporters of the plastid organelle in Apicomplexa.Curr Opin Microbiol. 2021 Oct;63:250-258. doi: 10.1016/j.mib.2021.07.016. Epub 2021 Aug 26. Curr Opin Microbiol. 2021. PMID: 34455306 Review.
-
Parasite powerhouse: A review of the Toxoplasma gondii mitochondrion.J Eukaryot Microbiol. 2022 Nov;69(6):e12906. doi: 10.1111/jeu.12906. Epub 2022 May 4. J Eukaryot Microbiol. 2022. PMID: 35315174 Free PMC article. Review.
Cited by
-
Autophagy-Related Protein ATG18 Regulates Apicoplast Biogenesis in Apicomplexan Parasites.mBio. 2017 Oct 31;8(5):e01468-17. doi: 10.1128/mBio.01468-17. mBio. 2017. PMID: 29089429 Free PMC article.
-
PI3-kinase has multiple functions in asexual blood stages of Plasmodium falciparum.Sci Rep. 2025 May 14;15(1):16762. doi: 10.1038/s41598-025-01397-1. Sci Rep. 2025. PMID: 40369090 Free PMC article.
-
Comparative Phosphoproteomic Analysis of Sporulated Oocysts and Tachyzoites of Toxoplasma gondii Reveals Stage-Specific Patterns.Molecules. 2022 Feb 2;27(3):1022. doi: 10.3390/molecules27031022. Molecules. 2022. PMID: 35164288 Free PMC article.
-
The conserved apicomplexan Aurora kinase TgArk3 is involved in endodyogeny, duplication rate and parasite virulence.Cell Microbiol. 2016 Aug;18(8):1106-1120. doi: 10.1111/cmi.12571. Epub 2016 Feb 22. Cell Microbiol. 2016. PMID: 26833682 Free PMC article.
-
A VPS15-like kinase regulates apicoplast biogenesis and autophagy by promoting PI3P generation in Toxoplasma gondii.PLoS Pathog. 2022 Nov 1;18(11):e1010922. doi: 10.1371/journal.ppat.1010922. eCollection 2022 Nov. PLoS Pathog. 2022. PMID: 36318587 Free PMC article.
References
-
- Achbarou A, Mercereau-Puijalon O, Autheman JM, Fortier B, Camus D, Dubremetz JF. Characterization of microneme proteins of Toxoplasma gondii. Mol Biochem Parasitol. 1991;47:223–233. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
