

The rv1184c locus encodes Chp2, an acyltransferase in Mycobacterium tuberculosis polyacyltrehalose lipid biosynthesis
- PMID: 25331437
- PMCID: PMC4288680
- DOI: 10.1128/JB.02015-14
The rv1184c locus encodes Chp2, an acyltransferase in Mycobacterium tuberculosis polyacyltrehalose lipid biosynthesis
Abstract
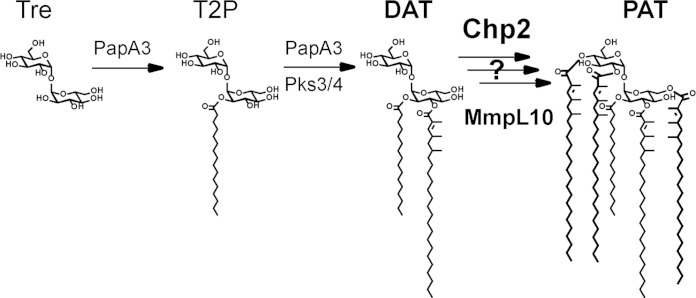
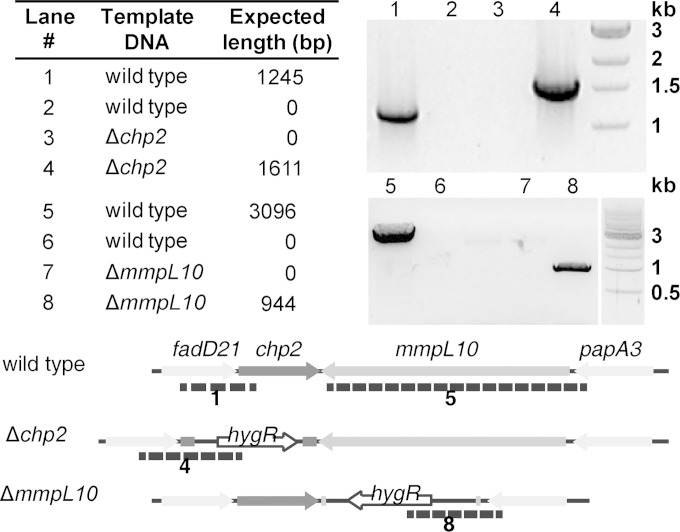
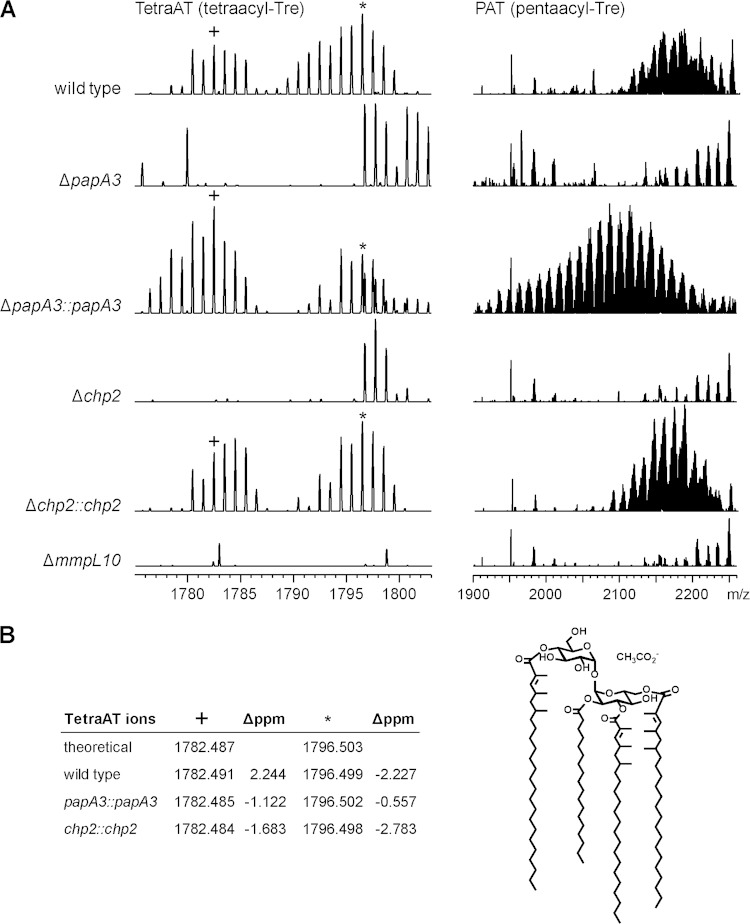
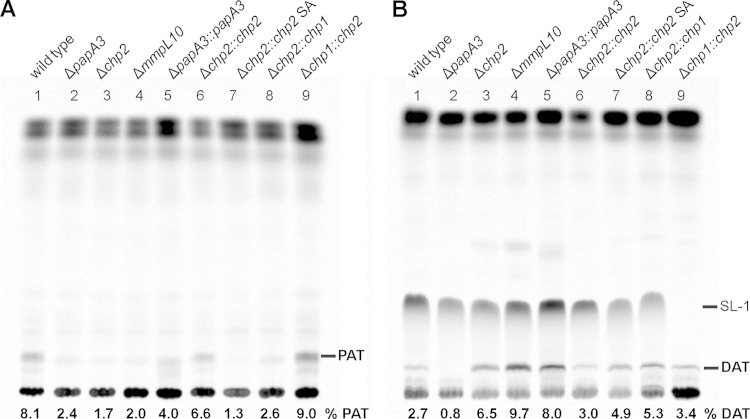
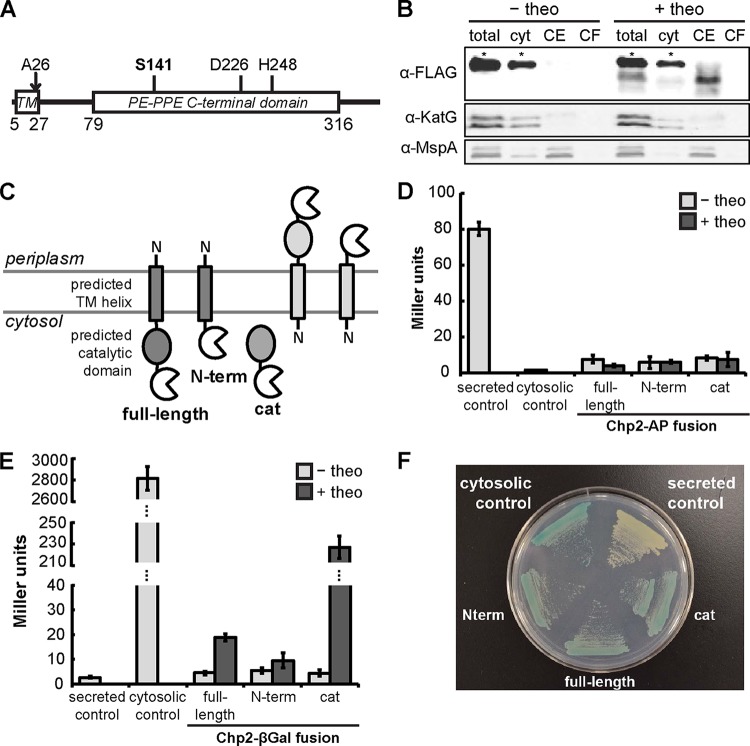
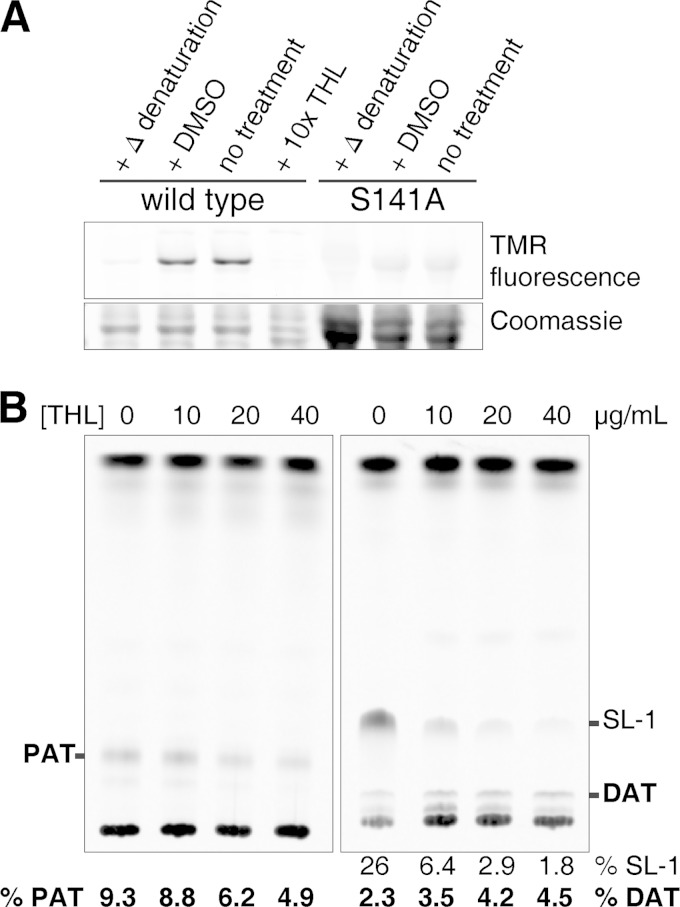
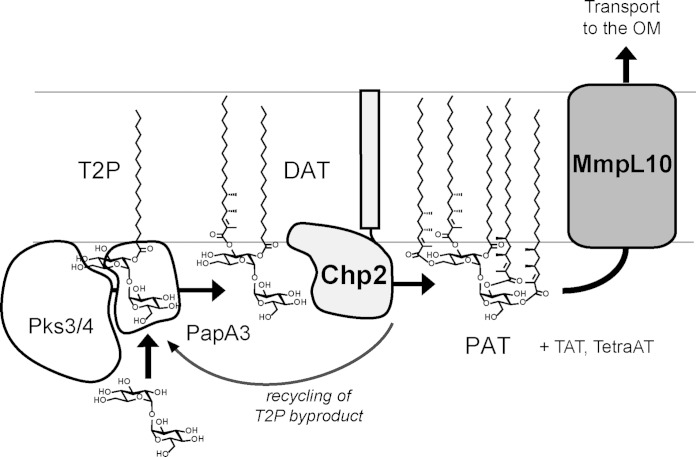
Trehalose glycolipids are found in many bacteria in the suborder Corynebacterineae, but methyl-branched acyltrehaloses are exclusive to virulent species such as the human pathogen Mycobacterium tuberculosis. In M. tuberculosis, the acyltransferase PapA3 catalyzes the formation of diacyltrehalose (DAT), but the enzymes responsible for downstream reactions leading to the final product, polyacyltrehalose (PAT), have not been identified. The PAT biosynthetic gene locus is similar to that of another trehalose glycolipid, sulfolipid 1. Recently, Chp1 was characterized as the terminal acyltransferase in sulfolipid 1 biosynthesis. Here we provide evidence that the homologue Chp2 (Rv1184c) is essential for the final steps of PAT biosynthesis. Disruption of chp2 led to the loss of PAT and a novel tetraacyltrehalose species, TetraAT, as well as the accumulation of DAT, implicating Chp2 as an acyltransferase downstream of PapA3. Disruption of the putative lipid transporter MmpL10 resulted in a similar phenotype. Chp2 activity thus appears to be regulated by MmpL10 in a relationship similar to that between Chp1 and MmpL8 in sulfolipid 1 biosynthesis. Chp2 is localized to the cell envelope fraction, consistent with its role in DAT modification and possible regulatory interactions with MmpL10. Labeling of purified Chp2 by an activity-based probe was dependent on the presence of the predicted catalytic residue Ser141 and was inhibited by the lipase inhibitor tetrahydrolipstatin (THL). THL treatment of M. tuberculosis resulted in selective inhibition of Chp2 over PapA3, confirming Chp2 as a member of the serine hydrolase superfamily. Efforts to produce in vitro reconstitution of acyltransferase activity using straight-chain analogues were unsuccessful, suggesting that Chp2 has specificity for native methyl-branched substrates.
Copyright © 2015, American Society for Microbiology. All Rights Reserved.
Figures
References
-
- Dubey VS, Sirakova TD, Kolattukudy PE. 2002. Disruption of msl3 abolishes the synthesis of mycolipanoic and mycolipenic acids required for polyacyltrehalose synthesis in Mycobacterium tuberculosis H37Rv and causes cell aggregation. Mol Microbiol 45:1451–1459. doi: 10.1046/j.1365-2958.2002.03119.x. - DOI - PubMed
-
- Rousseau C, Neyrolles O, Bordat Y, Giroux S, Sirakova TD, Prevost M-C, Kolattukudy PE, Gicquel B, Jackson M. 2003. Deficiency in mycolipenate- and mycosanoate-derived acyltrehaloses enhances early interactions of Mycobacterium tuberculosis with host cells. Cell Microbiol 5:405–415. doi: 10.1046/j.1462-5822.2003.00289.x. - DOI - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
