

Axl phosphorylates Elmo scaffold proteins to promote Rac activation and cell invasion
- PMID: 25332238
- PMCID: PMC4295393
- DOI: 10.1128/MCB.00764-14
Axl phosphorylates Elmo scaffold proteins to promote Rac activation and cell invasion
Abstract
The receptor tyrosine kinase Axl contributes to cell migration and invasion. Expression of Axl correlates with metastatic progression in cancer patients, yet the specific signaling events promoting invasion downstream of Axl are poorly defined. Herein, we report Elmo scaffolds to be direct substrates and binding partners of Axl. Elmo proteins are established to interact with Dock family guanine nucleotide exchange factors to control Rac-mediated cytoskeletal dynamics. Proteomics and mutagenesis studies reveal that Axl phosphorylates Elmo1/2 on a conserved carboxyl-terminal tyrosine residue. Upon Gas6-dependent activation of Axl, endogenous Elmo2 becomes phosphorylated on Tyr-713 and enters into a physical complex with Axl in breast cancer cells. Interfering with Elmo2 expression prevented Gas6-induced Rac1 activation in breast cancer cells. Similarly to blocking of Axl, Elmo2 knockdown or pharmacological inhibition of Dock1 abolishes breast cancer cell invasion. Interestingly, Axl or Elmo2 knockdown diminishes breast cancer cell proliferation. Rescue of Elmo2 knockdown cells with the wild-type protein but not with Elmo2 harboring Tyr-713-Phe mutations restores cell invasion and cell proliferation. These results define a new mechanism by which Axl promotes cell proliferation and invasion and identifies inhibition of the Elmo-Dock pathway as a potential therapeutic target to stop Axl-induced metastases.
Copyright © 2015, American Society for Microbiology. All Rights Reserved.
Figures
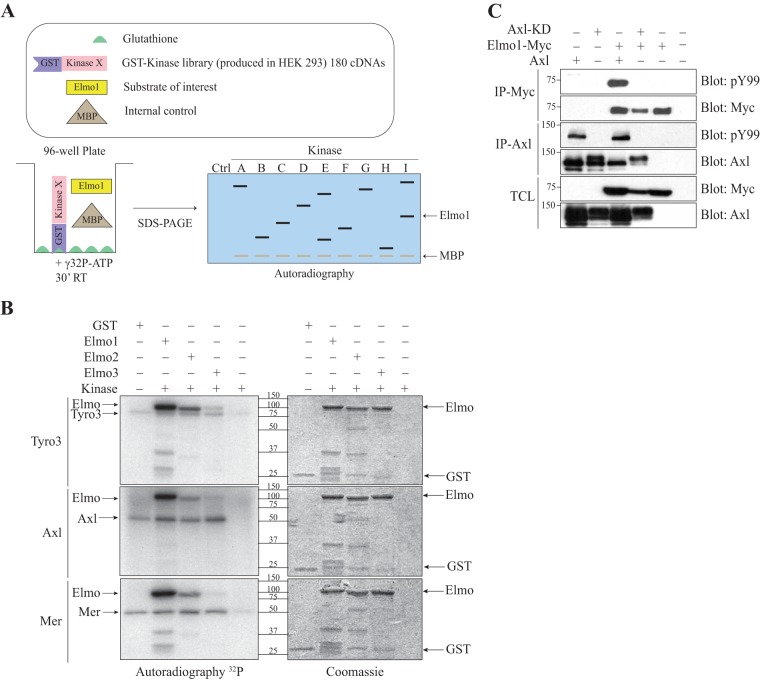
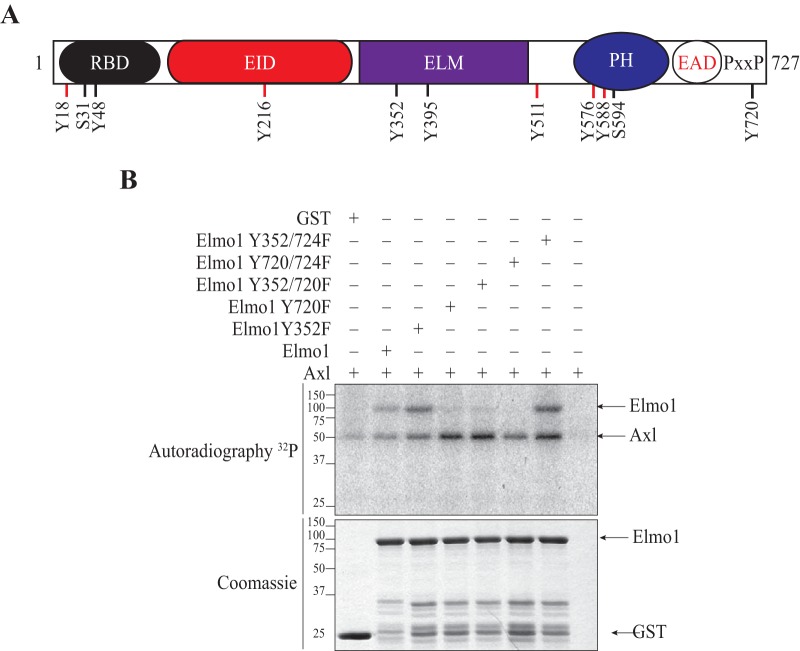
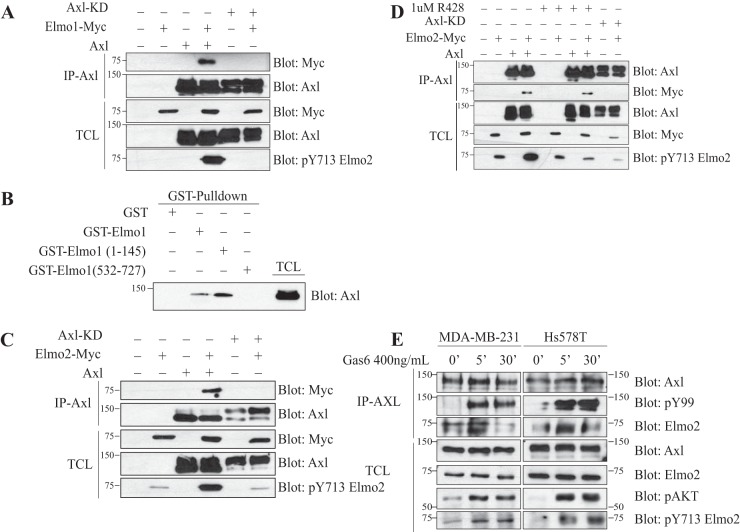
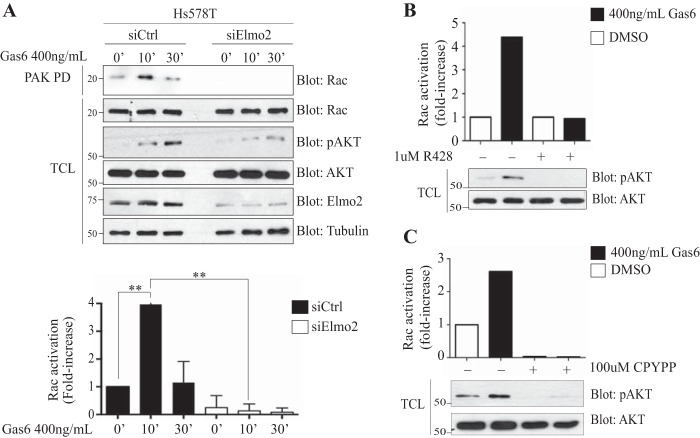
References
-
- Stitt TN, Conn G, Gore M, Lai C, Bruno J, Radziejewski C, Mattsson K, Fisher J, Gies DR, Jones PF, Masiakowski P, Ryan TE, Tobkes NJ, Chen DH, DiStefano PS, Long GL, Basilico C, Goldfarb MP, Lemke G, Glass DJ, Yancopoulos GD. 1995. The anticoagulation factor protein S and its relative, Gas6, are ligands for the Tyro 3/Axl family of receptor tyrosine kinases. Cell 80:661–670. doi:10.1016/0092-8674(95)90520-0. - DOI - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous