

Interleukin-6 mediated upregulation of CYP1B1 and CYP2E1 in colorectal cancer involves DNA methylation, miR27b and STAT3
- PMID: 25333344
- PMCID: PMC4264448
- DOI: 10.1038/bjc.2014.540
Interleukin-6 mediated upregulation of CYP1B1 and CYP2E1 in colorectal cancer involves DNA methylation, miR27b and STAT3
Abstract
Background: The pro-inflammatory cytokine interleukin-6 (IL6) promotes colorectal cancer (CRC) development. It is also known to regulate cytochrome P450 (CYP450) enzymes, which are involved in CRC tumour initiation and promotion via activation of chemical carcinogens. Here, IL6 regulation of CYP450 expression was investigated in CRC.
Methods: The effect of IL6 on CYP 1A1, 1B1 and 2E1 expression was determined in vitro using CRC cell lines HCT116 and SW480, and CYP450 expression was determined by immunohistochemistry in CRC tissues previously shown to have increased levels of IL6.
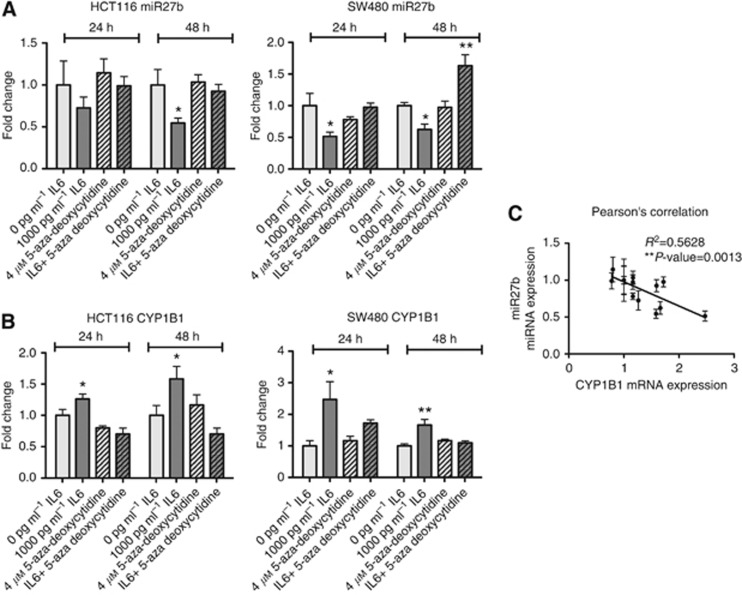
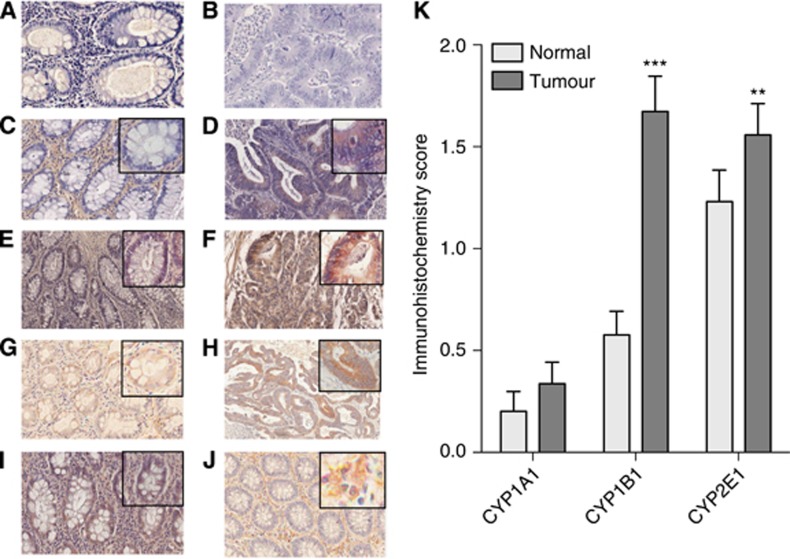
Results: In mechanistic studies, IL6 treatment significantly induced CYP1B1 and CYP2E1, but not CYP1A1, gene expression in HCT116 and SW480 cells. CYP2E1 expression regulation occurred via a transcriptional mechanism involving STAT3. For CYP1B1 regulation, IL6 downregulated the CYP1B1-targeting microRNA miR27b through a mechanism involving DNA methylation. In clinical samples, the expression of CYP1B1 and CYP2E1, but not CYP1A1, was significantly increased in malignant tissue overexpressing IL6 compared with matched adjacent normal tissue.
Conclusions: Colonic inflammation with the presence of IL6 associated with neoplastic tissue can alter metabolic competency of epithelial cells by manipulating CYP2E1 and CYP1B1 expression through transcriptional and epigenetic mechanisms. This can lead to increased activation of dietary carcinogens and DNA damage, thus promoting colorectal carcinogenesis.
Figures





References
-
- Abdel-Razzak Z, Loyer P, Fautrel A, Gautier JC, Corcos L, Turlin B, Beaune P, Guillouzo A. Cytokines down-regulate expression of major cytochrome P-450 enzymes in adult human hepatocytes in primary culture. Mol Pharmacol. 1993;44:707–715. - PubMed
-
- Barrett JH, Smith G, Waxman R, Gooderham N, Lightfoot T, Garner RC, Augustsson K, Wolf CR, Bishop DT, Forman D, Colorectal Cancer Study Group Investigation of interaction between N-acetyltransferase 2 and heterocyclic amines as potential risk factors for colorectal cancer. Carcinogenesis. 2003;24:275–282. - PubMed
-
- Boyce A, Doehmer J, Gooderham NJ. Phytoalexin resveratrol attenuates the mutagenicity of the heterocyclic amines 2-amino-1-methyl-6-phenylimidazo[4,5-b]pyridine and 2-amino-3,8-dimethylimidazo[4,5-f]quinoxaline. J Chromatogr B Analyt Technol Biomed Life Sci. 2004;802 (1:217–223. - PubMed
-
- Braeuning A, Kohle C, Buchmann A, Schwarz M. Coordinate regulation of cytochrome P450 1a1 expression in mouse liver by the aryl hydrocarbon receptor and the beta-catenin pathway. Toxicol Sci. 2011;122:16–25. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
