

Nasal immunity is an ancient arm of the mucosal immune system of vertebrates
- PMID: 25335508
- PMCID: PMC4321879
- DOI: 10.1038/ncomms6205
Nasal immunity is an ancient arm of the mucosal immune system of vertebrates
Abstract
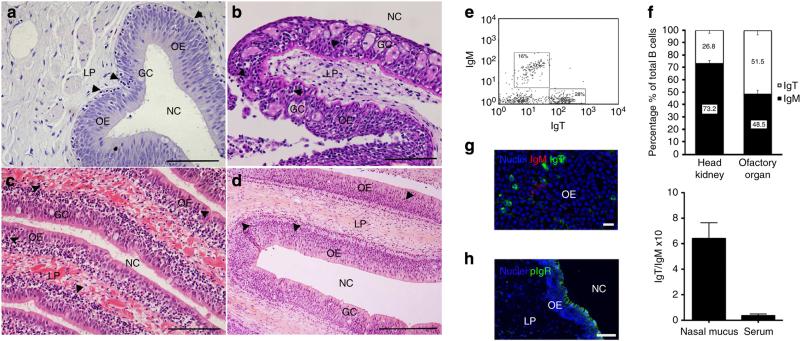
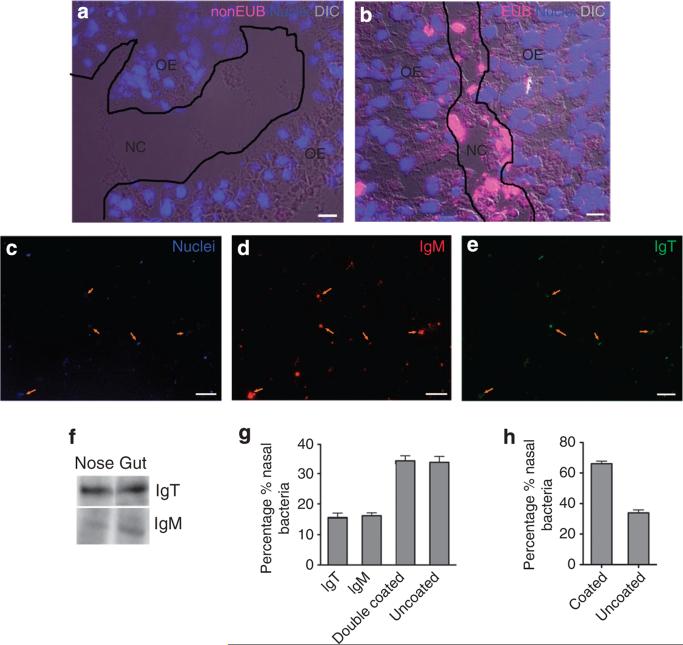
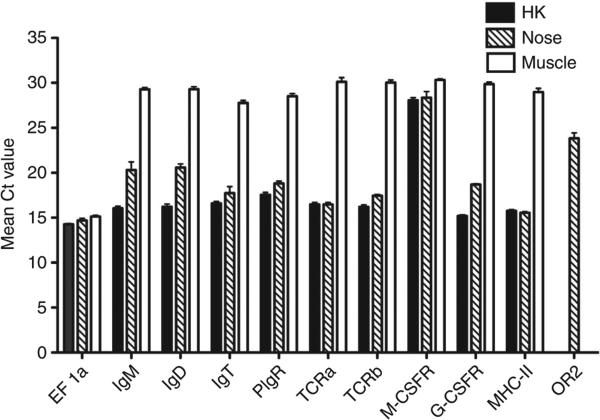
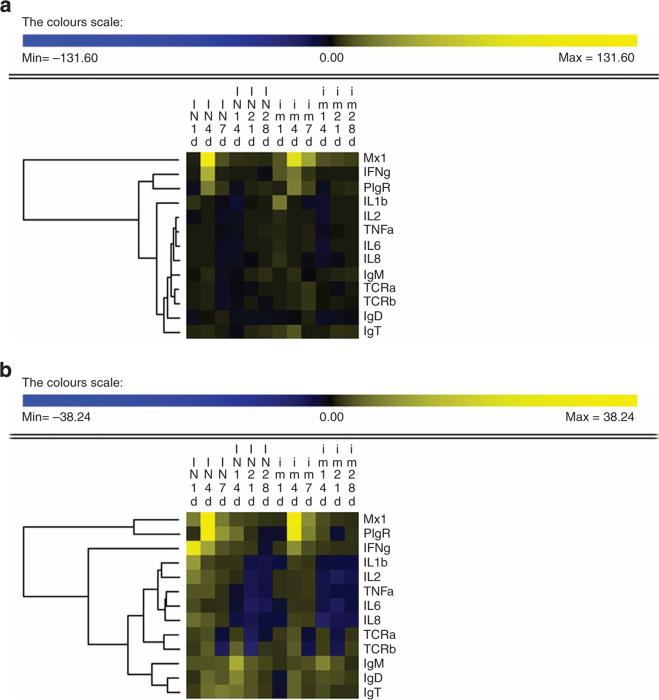
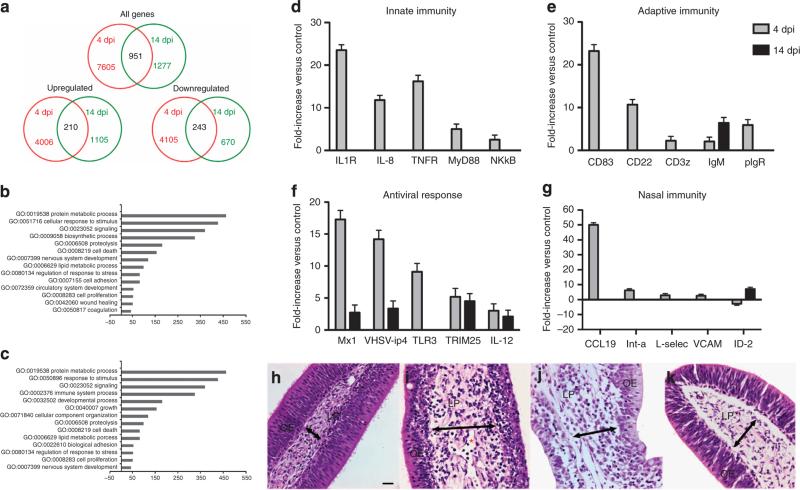
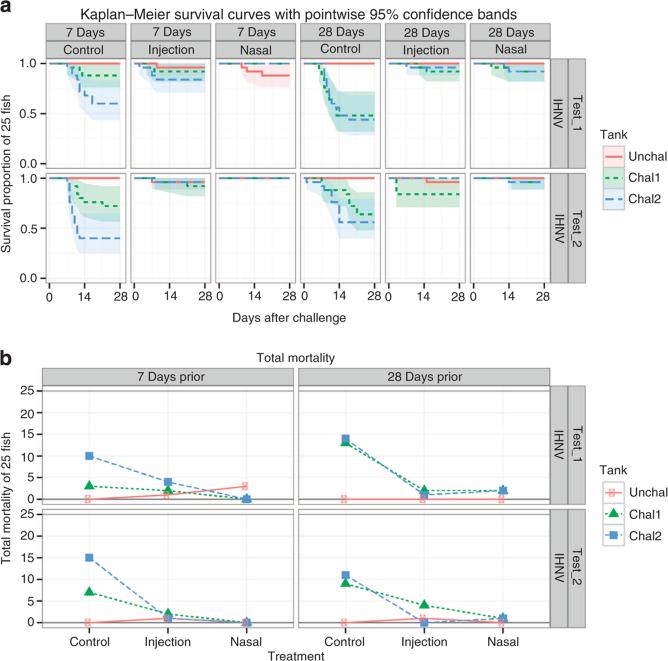
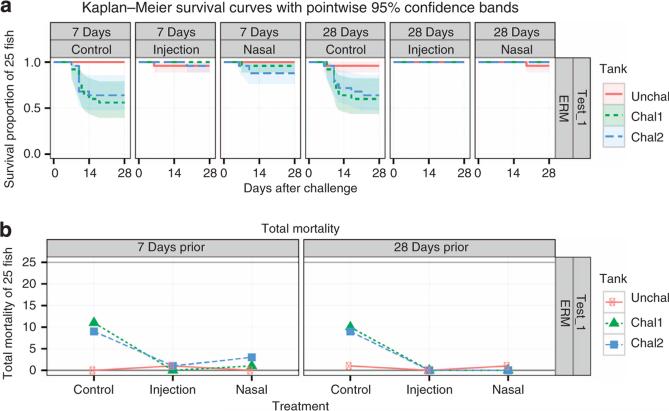
The mucosal surfaces of all vertebrates have been exposed to similar evolutionary pressures for millions of years. In terrestrial vertebrates such as birds and mammals, the nasopharynx-associated lymphoid tissue (NALT) represents a first line of immune defence. Here we propose that NALT is an ancient arm of the mucosal immune system not restricted to terrestrial vertebrates. We find that NALT is present in rainbow trout and that it resembles other teleost mucosa-associated lymphoid tissues. Trout NALT consists of diffuse lymphoid cells and lacks tonsils and adenoids. The predominant B-cell subset found in trout NALT are IgT(+) B cells, similar to skin and gut. The trout olfactory organ is colonized by abundant symbiotic bacteria, which are coated by trout secretory immunoglobulin. Trout NALT is capable of mounting strong anti-viral immune responses following nasal delivery of a live attenuated viral vaccine. Our results open up a new tool for the control of aquatic infectious diseases via nasal vaccination.
Figures
References
-
- Ache BW, Young JM. Olfaction: diverse species, conserved principles. Neuron. 2005;48:417–430. - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
