

Cellular microRNAs up-regulate transcription via interaction with promoter TATA-box motifs
- PMID: 25336585
- PMCID: PMC4238354
- DOI: 10.1261/rna.045633.114
Cellular microRNAs up-regulate transcription via interaction with promoter TATA-box motifs
Abstract
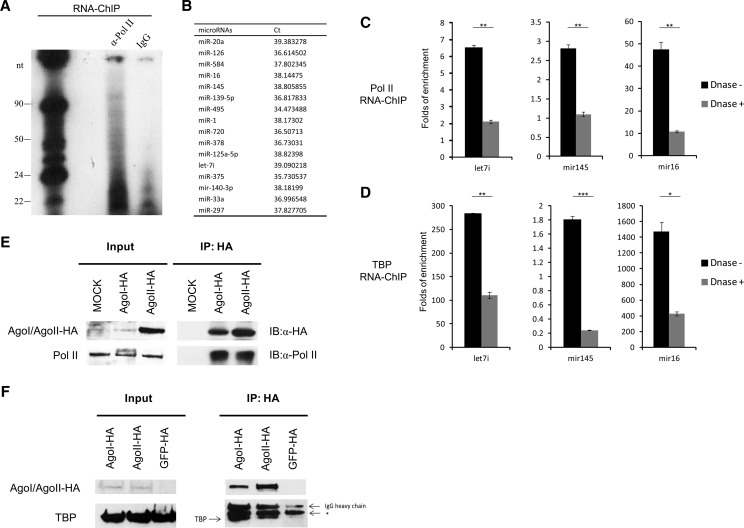
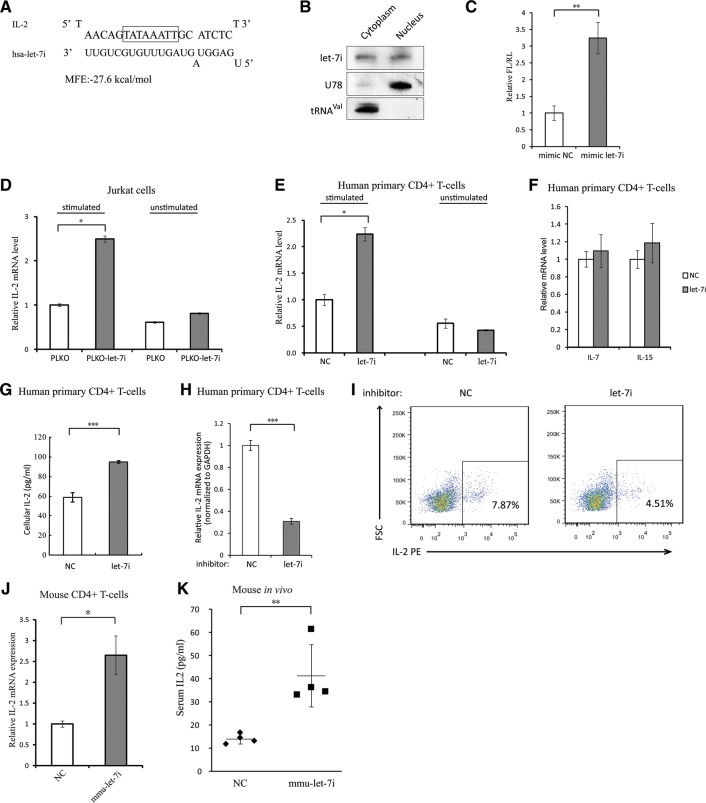
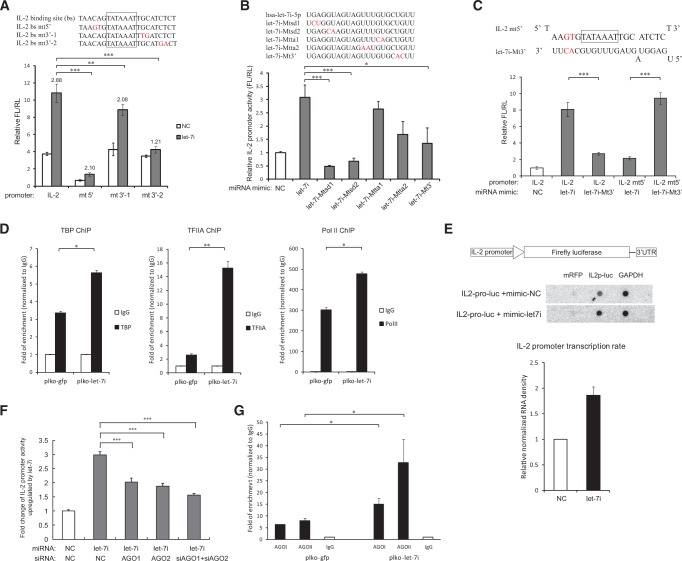
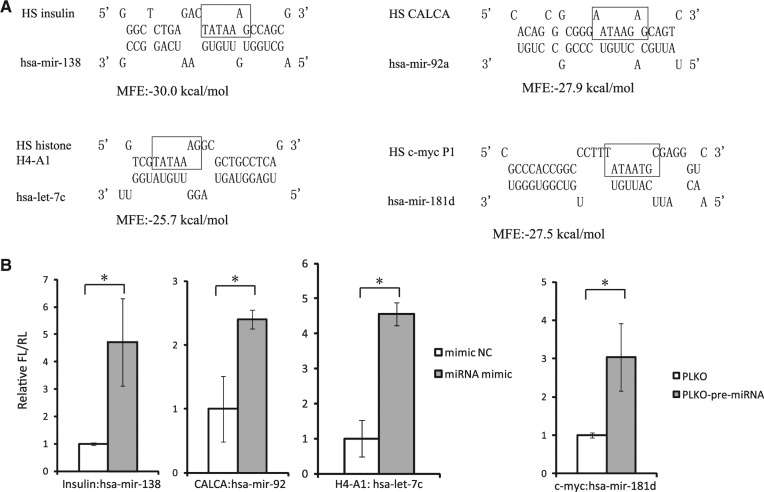
The TATA box represents one of the most prevalent core promoters where the pre-initiation complexes (PICs) for gene transcription are assembled. This assembly is crucial for transcription initiation and well regulated. Here we show that some cellular microRNAs (miRNAs) are associated with RNA polymerase II (Pol II) and TATA box-binding protein (TBP) in human peripheral blood mononuclear cells (PBMCs). Among them, let-7i sequence specifically binds to the TATA-box motif of interleukin-2 (IL-2) gene and elevates IL-2 mRNA and protein production in CD4(+) T-lymphocytes in vitro and in vivo. Through direct interaction with the TATA-box motif, let-7i facilitates the PIC assembly and transcription initiation of IL-2 promoter. Several other cellular miRNAs, such as mir-138, mir-92a or mir-181d, also enhance the promoter activities via binding to the TATA-box motifs of insulin, calcitonin or c-myc, respectively. In agreement with the finding that an HIV-1-encoded miRNA could enhance viral replication through targeting the viral promoter TATA-box motif, our data demonstrate that the interaction with core transcription machinery is a novel mechanism for miRNAs to regulate gene expression.
Keywords: TATA-box motif; core transcription machinery; microRNAs; transcription activation.
© 2014 Zhang et al.; Published by Cold Spring Harbor Laboratory Press for the RNA Society.
Figures
References
-
- Bartel DP 2004. MicroRNAs: genomics, biogenesis, mechanism, and function. Cell 116: 281–297. - PubMed
-
- Burley SK, Roeder RG 1996. Biochemistry and structural biology of transcription factor IID (TFIID). Annu Rev Biochem 65: 769–799. - PubMed
-
- Bushati N, Cohen SM 2007. microRNA functions. Annu Rev Cell Dev Biol 23: 175–205. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials