

MONOPTEROS directly activates the auxin-inducible promoter of the Dof5.8 transcription factor gene in Arabidopsis thaliana leaf provascular cells
- PMID: 25336688
- PMCID: PMC4265163
- DOI: 10.1093/jxb/eru418
MONOPTEROS directly activates the auxin-inducible promoter of the Dof5.8 transcription factor gene in Arabidopsis thaliana leaf provascular cells
Abstract
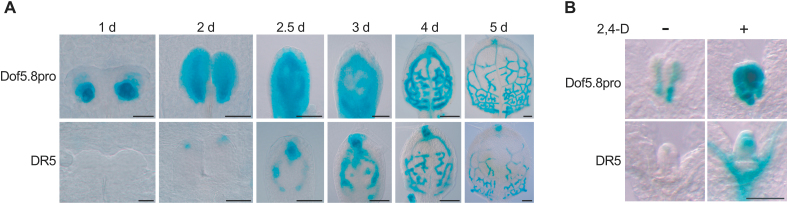
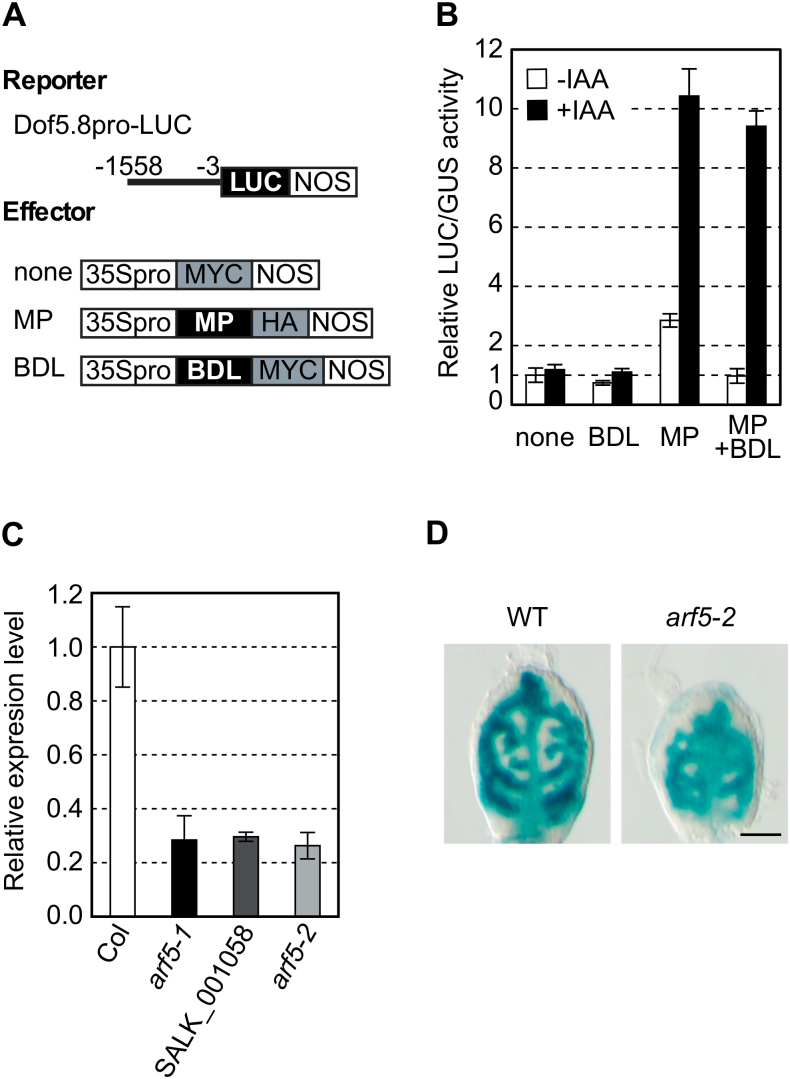
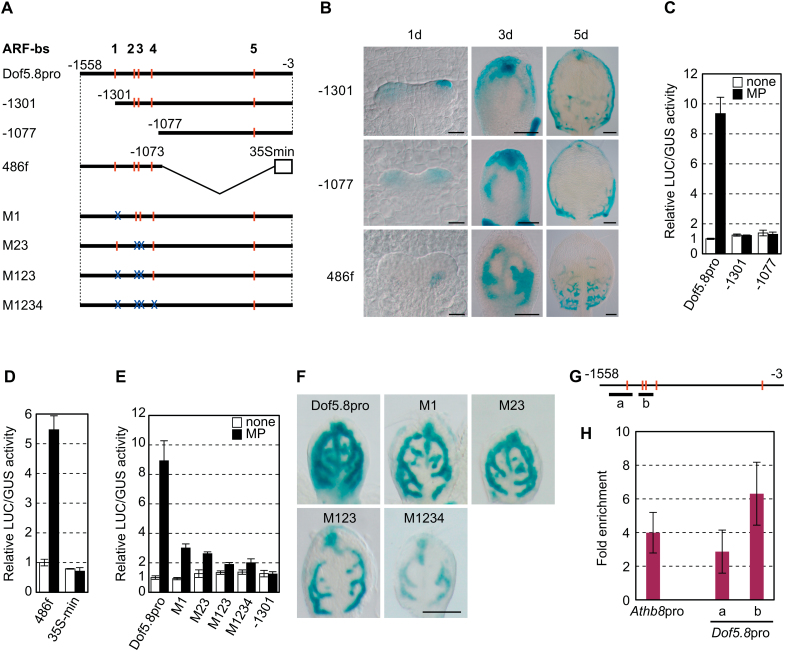
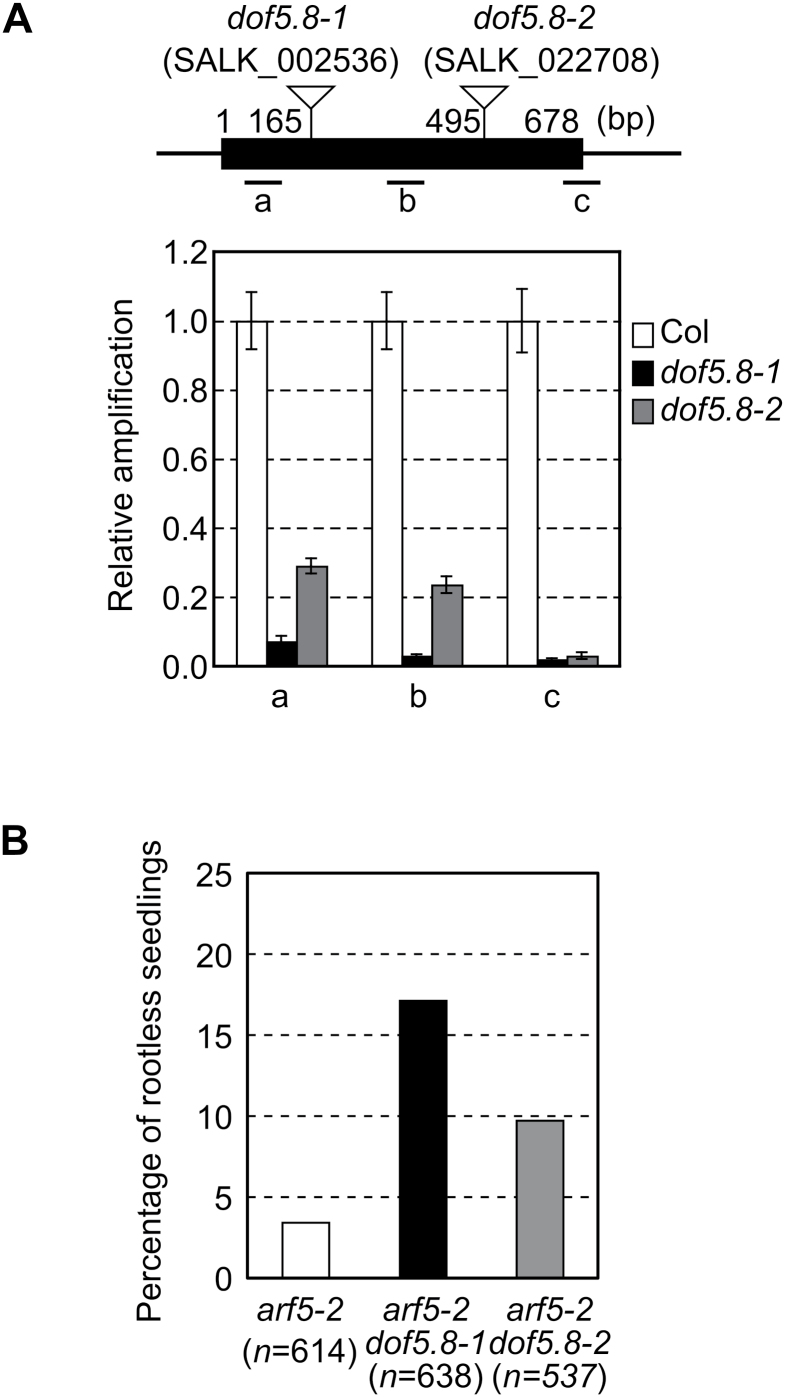
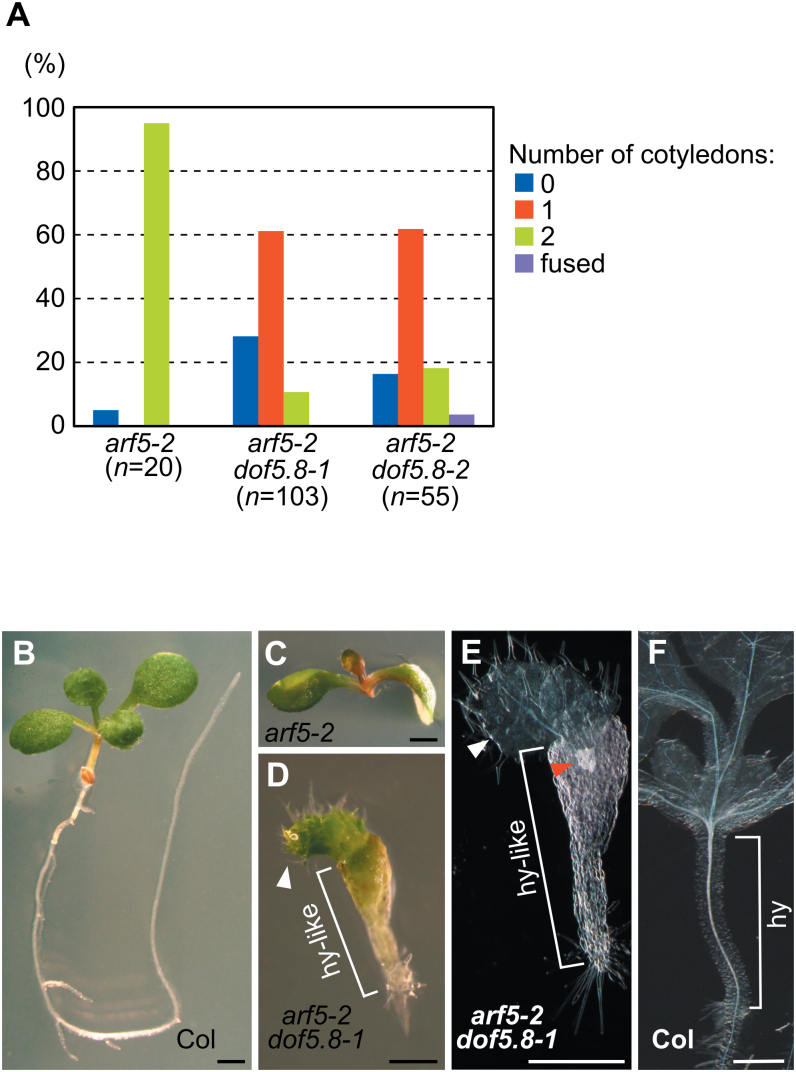
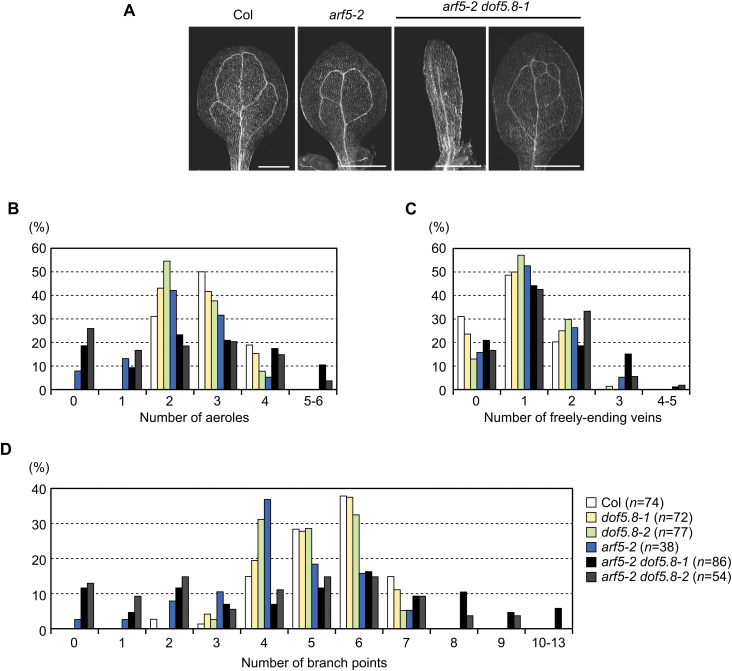
MONOPTEROS (MP) is an auxin-responsive transcription factor that is required for primary root formation and vascular development, whereas Dof5.8 is a Dof-class transcription factor whose gene is expressed in embryos as well as the pre- and procambial cells in the leaf primordium in Arabidopsis thaliana. In this study, it is shown that MP directly activates the Dof5.8 promoter. Although no apparent phenotype of the single dof5.8 mutants was found, phenotypic analysis with the mp dof5.8 double mutants revealed that mutations within Dof5.8 enhanced the phenotype of a weak allele of mp, with an increase in the penetrance of the 'rootless' phenotype and a reduction in the number of cotyledons. Furthermore, interestingly, although mp mutants showed reduced vascular pattern complexity in cotyledons, the mp dof5.8 double mutants displayed both more simplex and more complex vascular patterns in individual cotyledons. These results imply that the product of Dof5.8 whose expression is regulated by MP at least in part might be involved in multiple processes controlled by MP.
Keywords: Arabidopsis thaliana; Dof transcription factor; MONOPTEROS; auxin response; embryo development; vascular development..
© The Author 2014. Published by Oxford University Press on behalf of the Society for Experimental Biology.
Figures
References
-
- Alonso JM, Stepanova AN, Leisse TJ, et al. 2003. Genome-wide insertional mutagenesis of Arabidopsis thaliana . Science 301, 653–657. - PubMed
-
- Bauby H, Divol F, Truernit E, Grandjean O, Palauqui JC. 2007. Protophloem differentiation in early Arabidopsis thaliana development. Plant and Cell Physiology 48, 97–109. - PubMed
-
- Berleth T, Jürgens G. 1993. The role of the monopteros gene in organising the basal body region of the Arabidopsis embryo. Development 118, 575–587.
-
- Beuchat J, Scacchi E, Tarkowska D, Ragni L, Strnad M, Hardtke CS. 2010. BRX promotes Arabidopsis shoot growth. New Phytologist 188, 23–29. - PubMed
-
- Ckurshumova W, Scarpella E, Goldstein RS, Berleth T. 2011. Double-filter identification of vascular-expressed genes using Arabidopsis plants with vascular hypertrophy and hypotrophy. Plant Science 181, 96–104. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
