

Broccoli: rapid selection of an RNA mimic of green fluorescent protein by fluorescence-based selection and directed evolution
- PMID: 25337688
- PMCID: PMC4244833
- DOI: 10.1021/ja508478x
Broccoli: rapid selection of an RNA mimic of green fluorescent protein by fluorescence-based selection and directed evolution
Abstract
Genetically encoded fluorescent ribonucleic acids (RNAs) have diverse applications, including imaging RNA trafficking and as a component of RNA-based sensors that exhibit fluorescence upon binding small molecules in live cells. These RNAs include the Spinach and Spinach2 aptamers, which bind and activate the fluorescence of fluorophores similar to that found in green fluorescent protein. Although additional highly fluorescent RNA-fluorophore complexes would extend the utility of this technology, the identification of novel RNA-fluorophore complexes is difficult. Current approaches select aptamers on the basis of their ability to bind fluorophores, even though fluorophore binding alone is not sufficient to activate fluorescence. Additionally, aptamers require extensive mutagenesis to efficiently fold and exhibit fluorescence in living cells. Here we describe a platform for rapid generation of highly fluorescent RNA-fluorophore complexes that are optimized for function in cells. This procedure involves selection of aptamers on the basis of their binding to fluorophores, coupled with fluorescence-activated cell sorting (FACS) of millions of aptamers expressed in Escherichia coli. Promising aptamers are then further optimized using a FACS-based directed evolution approach. Using this approach, we identified several novel aptamers, including a 49-nt aptamer, Broccoli. Broccoli binds and activates the fluorescence of (Z)-4-(3,5-difluoro-4-hydroxybenzylidene)-1,2-dimethyl-1H-imidazol-5(4H)-one. Broccoli shows robust folding and green fluorescence in cells, and increased fluorescence relative to Spinach2. This reflects, in part, improved folding in the presence of low cytosolic magnesium concentrations. Thus, this novel fluorescence-based selection approach simplifies the generation of aptamers that are optimized for expression and performance in living cells.
Figures
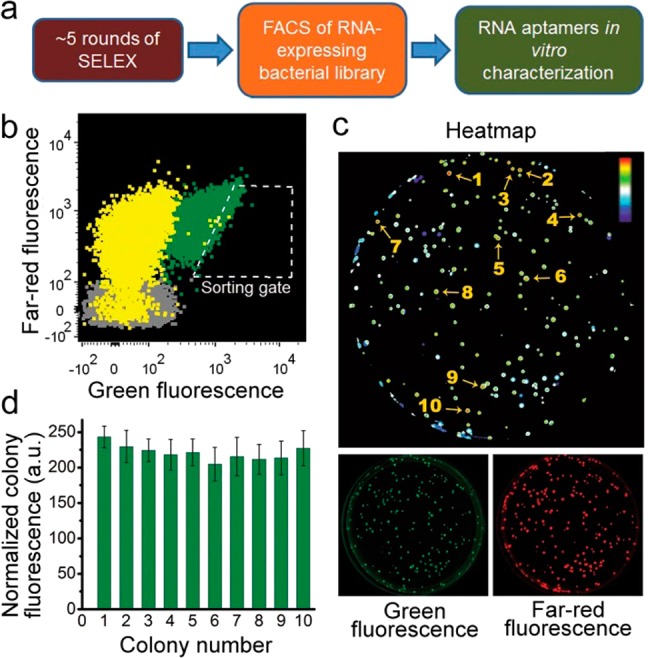


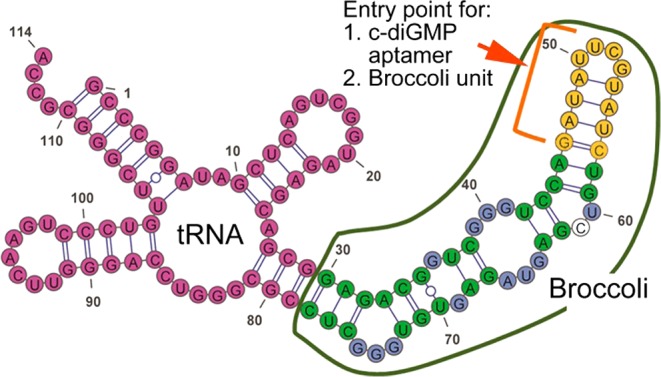
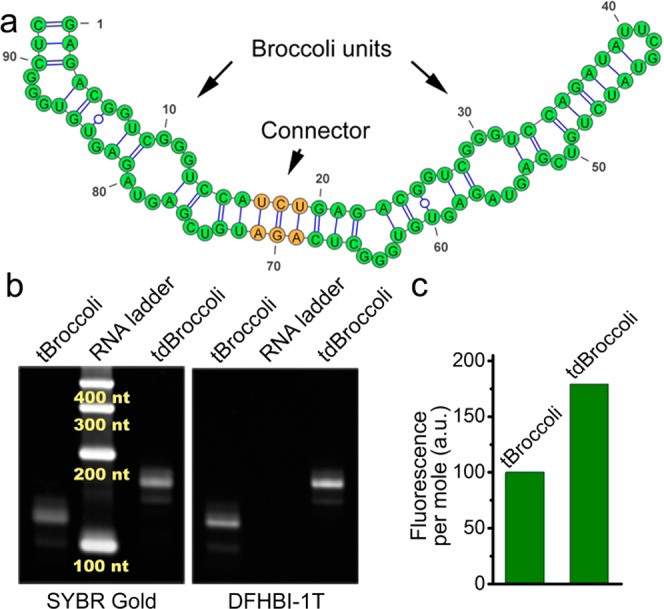

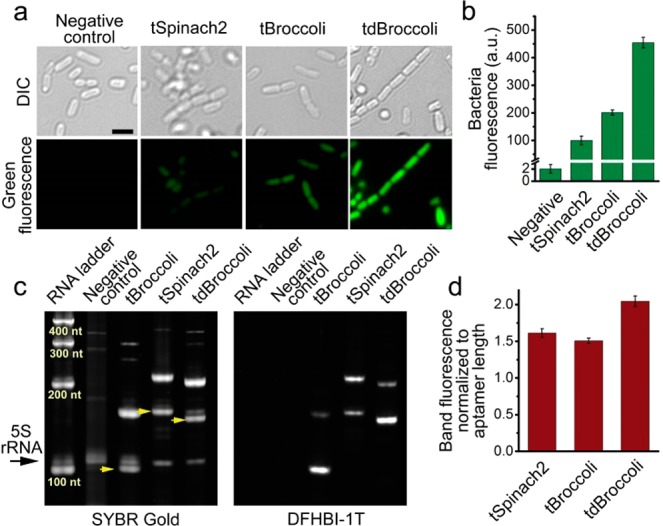

References
-
- Tyagi S. Nat. Methods 2009, 6, 331. - PubMed
-
- Bertrand E.; Chartrand P.; Schaefer M.; Shenoy S. M.; Singer R. H.; Long R. M. Mol. Cell 1998, 2, 437. - PubMed
-
- Babendure J. R.; Adams S. R.; Tsien R. Y. J. Am. Chem. Soc. 2003, 125, 14716. - PubMed
-
- Constantin T. P.; Silva G. L.; Robertson K. L.; Hamilton T. P.; Fague K.; Waggoner A. S.; Armitage B. A. Org. Lett. 2008, 10, 1561. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
