

Loss of mitochondrial fission depletes axonal mitochondria in midbrain dopamine neurons
- PMID: 25339743
- PMCID: PMC4205554
- DOI: 10.1523/JNEUROSCI.0930-14.2014
Loss of mitochondrial fission depletes axonal mitochondria in midbrain dopamine neurons
Abstract
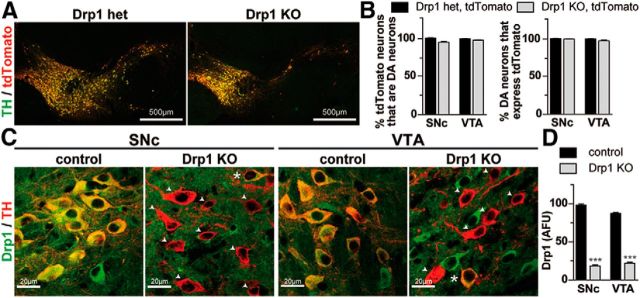
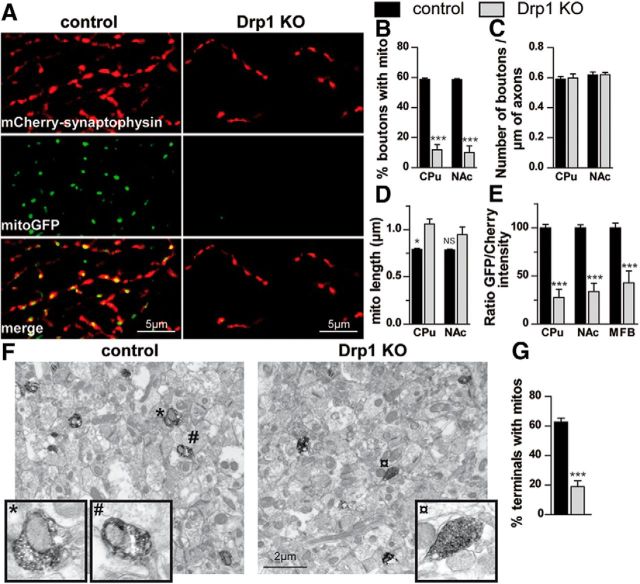
Disruptions in mitochondrial dynamics may contribute to the selective degeneration of dopamine (DA) neurons in Parkinson's disease (PD). However, little is known about the normal functions of mitochondrial dynamics in these neurons, especially in axons where degeneration begins, and this makes it difficult to understand the disease process. To study one aspect of mitochondrial dynamics-mitochondrial fission-in mouse DA neurons, we deleted the central fission protein dynamin-related protein 1 (Drp1). Drp1 loss rapidly eliminates the DA terminals in the caudate-putamen and causes cell bodies in the midbrain to degenerate and lose α-synuclein. Without Drp1, mitochondrial mass dramatically decreases, especially in axons, where the mitochondrial movement becomes uncoordinated. However, in the ventral tegmental area (VTA), a subset of midbrain DA neurons characterized by small hyperpolarization-activated cation currents (Ih) is spared, despite near complete loss of their axonal mitochondria. Drp1 is thus critical for targeting mitochondria to the nerve terminal, and a disruption in mitochondrial fission can contribute to the preferential death of nigrostriatal DA neurons.
Keywords: Drp1; Parkinson's disease; axon; mitochondria; neurodegeneration.
Copyright © 2014 the authors 0270-6474/14/3414304-14$15.00/0.
Figures









References
-
- Alexander C, Votruba M, Pesch UE, Thiselton DL, Mayer S, Moore A, Rodriguez M, Kellner U, Leo-Kottler B, Auburger G, Bhattacharya SS, Wissinger B. OPA1, encoding a dynamin-related GTPase, is mutated in autosomal dominant optic atrophy linked to chromosome 3q28. Nat Genet. 2000;26:211–215. doi: 10.1038/79944. - DOI - PubMed
-
- Arnoult D, Rismanchi N, Grodet A, Roberts RG, Seeburg DP, Estaquier J, Sheng M, Blackstone C. Bax/Bak-dependent release of DDP/TIMM8a promotes Drp1-mediated mitochondrial fission and mitoptosis during programmed cell death. Curr Biol. 2005;15:2112–2118. doi: 10.1016/j.cub.2005.10.041. - DOI - PubMed
-
- Berthet A, Porras G, Doudnikoff E, Stark H, Cador M, Bezard E, Bloch B. Pharmacological analysis demonstrates dramatic alteration of D1 dopamine receptor neuronal distribution in the rat analog of L-DOPA-induced dyskinesia. J Neurosci. 2009;29:4829–4835. doi: 10.1523/JNEUROSCI.5884-08.2009. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- P30NS069496/NS/NINDS NIH HHS/United States
- I01 BX001108/BX/BLRD VA/United States
- P30 NS065780/NS/NINDS NIH HHS/United States
- KO8NS062954/NS/NINDS NIH HHS/United States
- Wellcome Trust/United Kingdom
- OD010927/OD/NIH HHS/United States
- R01DA030529/DA/NIDA NIH HHS/United States
- P30NS065780/NS/NINDS NIH HHS/United States
- R01 DA030529/DA/NIDA NIH HHS/United States
- P30 NS069496/NS/NINDS NIH HHS/United States
- K26 OD010927/OD/NIH HHS/United States
- K08 NS062954/NS/NINDS NIH HHS/United States
- C06 RR018928/RR/NCRR NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous