

Dopamine invigorates reward seeking by promoting cue-evoked excitation in the nucleus accumbens
- PMID: 25339748
- PMCID: PMC4205557
- DOI: 10.1523/JNEUROSCI.3492-14.2014
Dopamine invigorates reward seeking by promoting cue-evoked excitation in the nucleus accumbens
Abstract
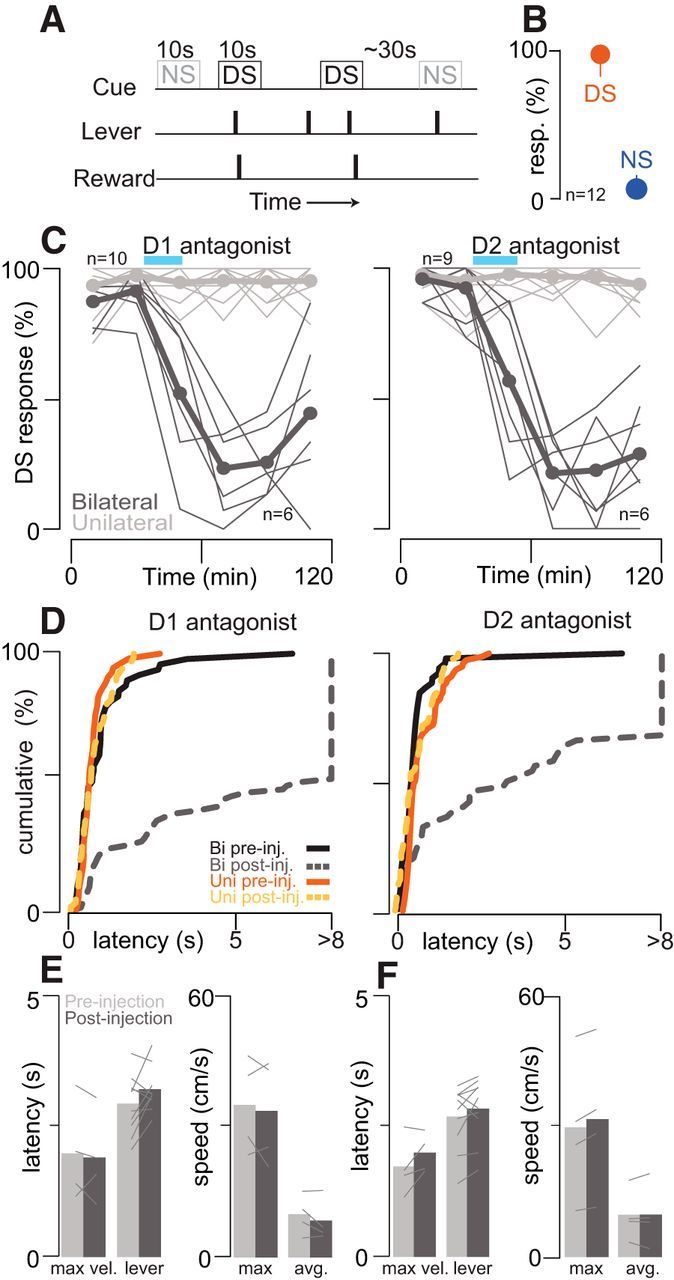
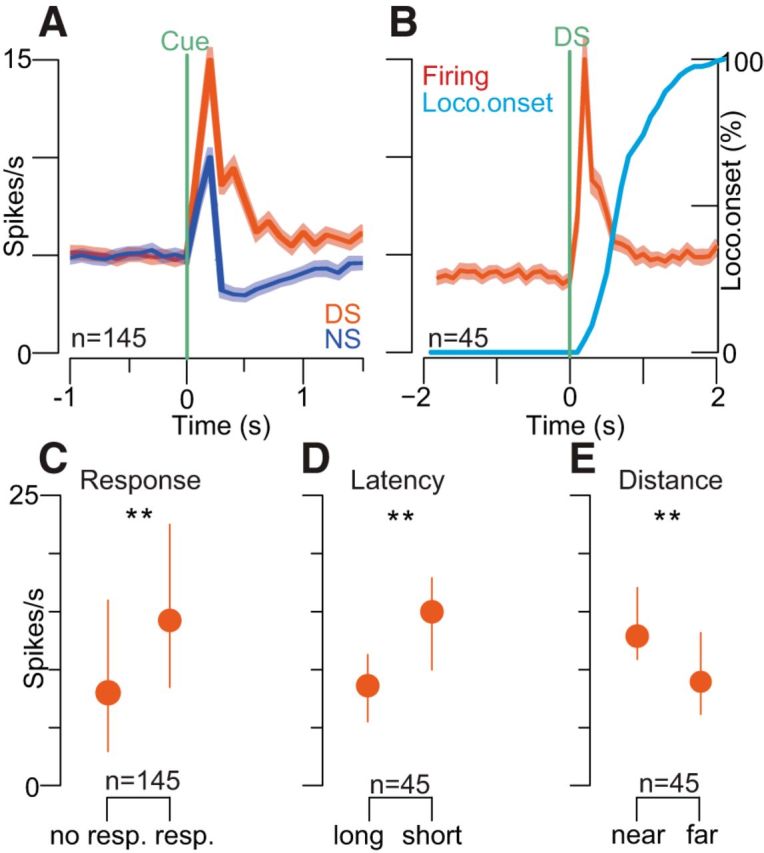
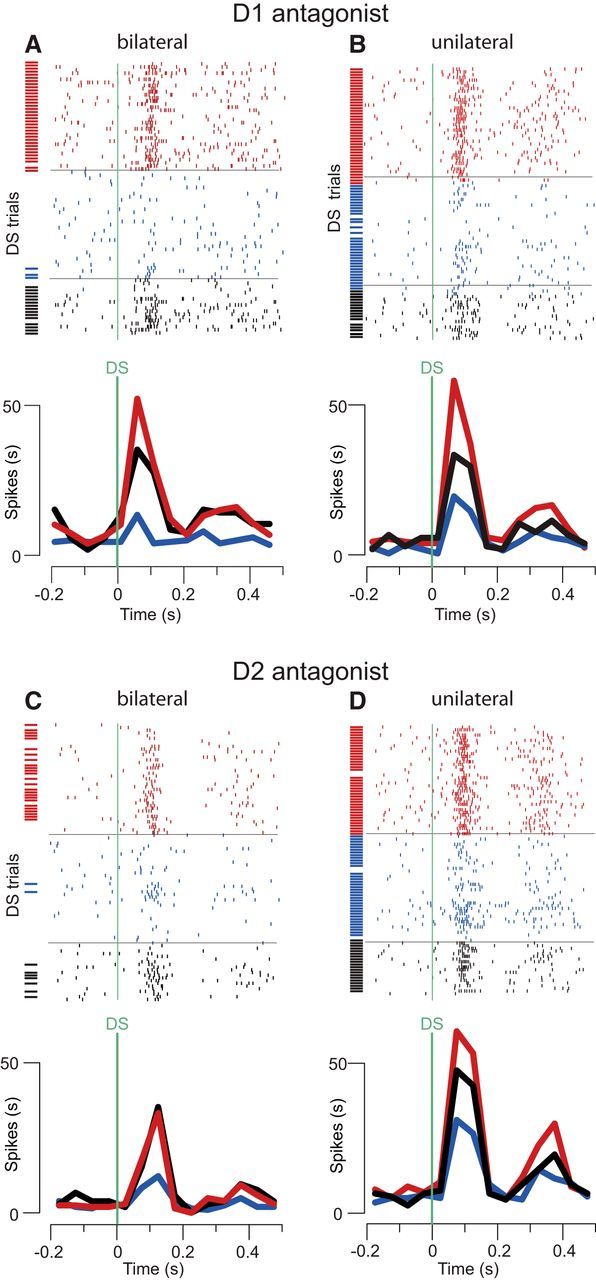
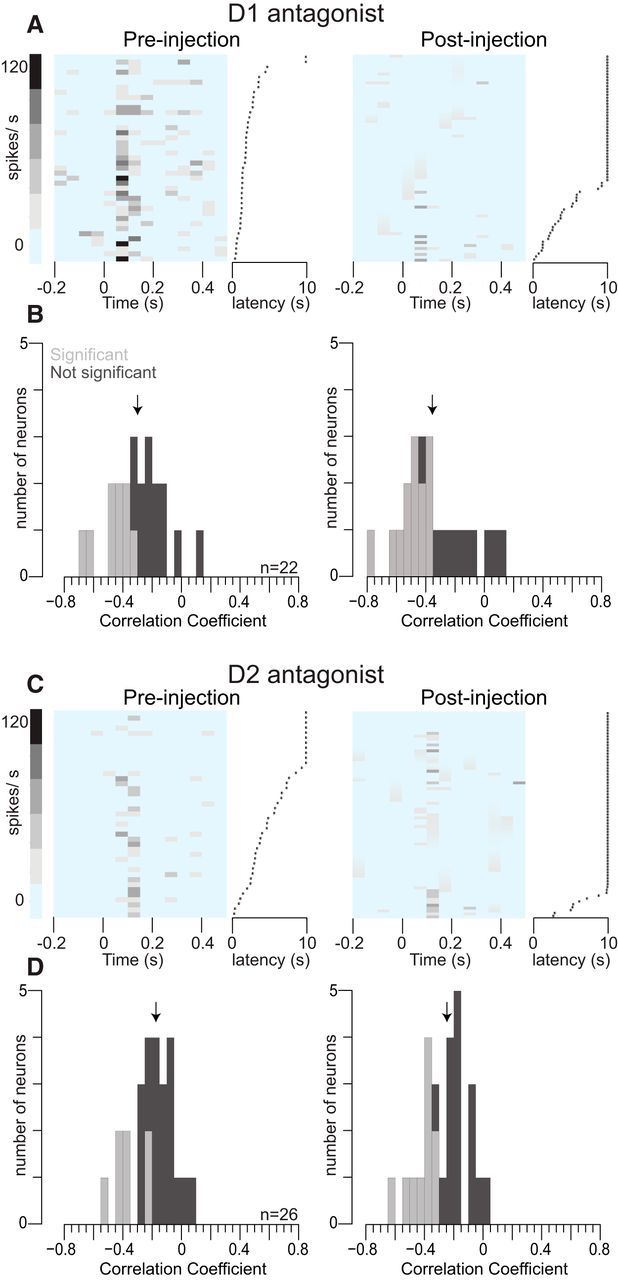
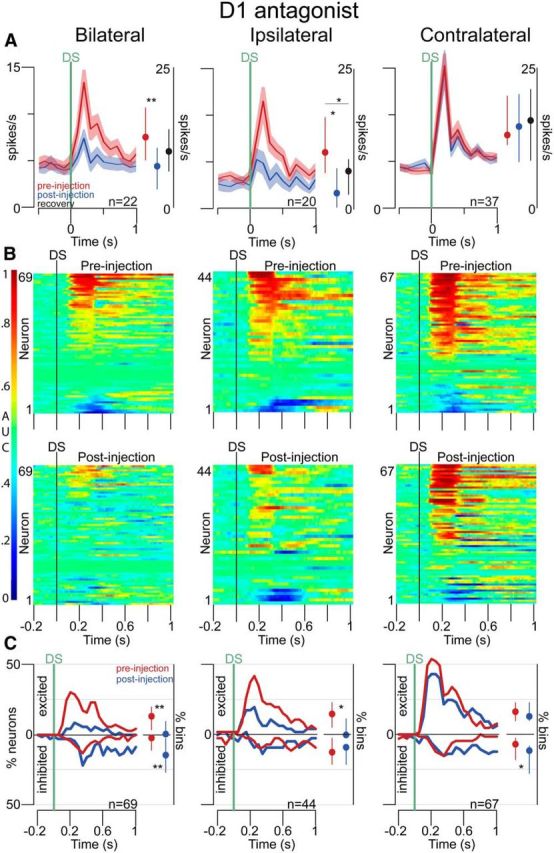
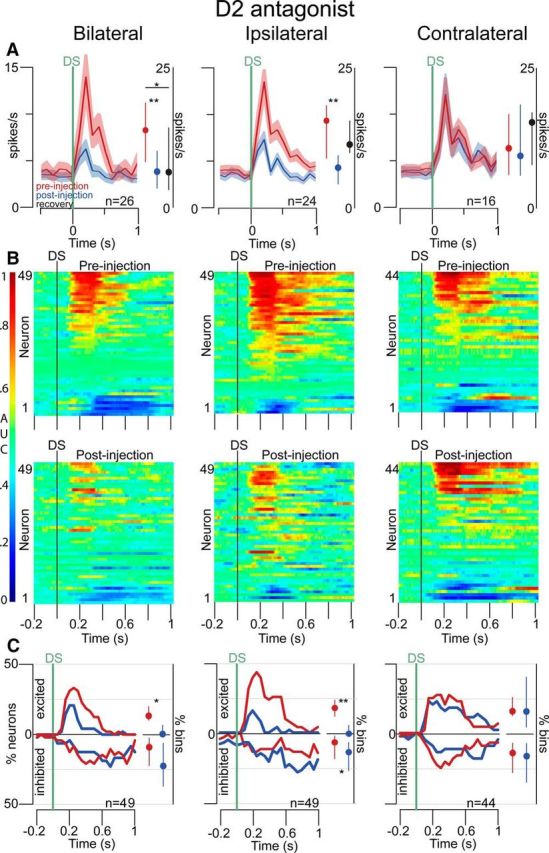
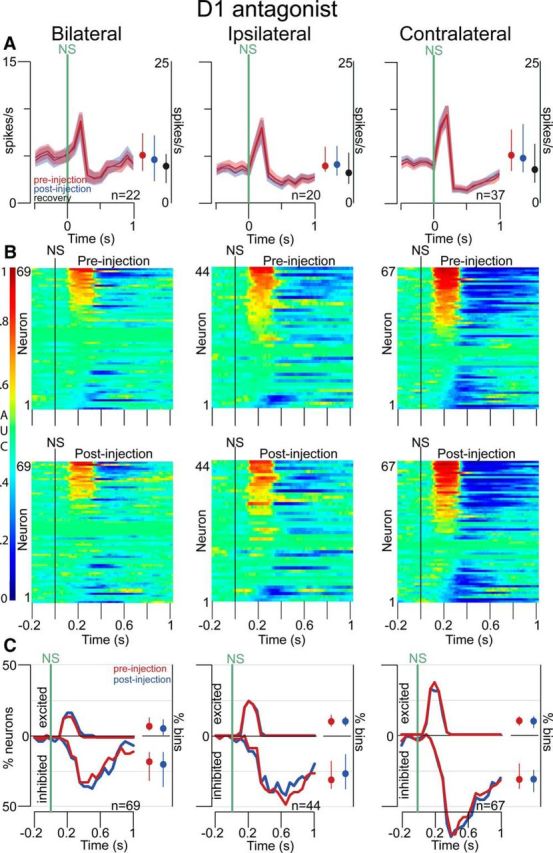
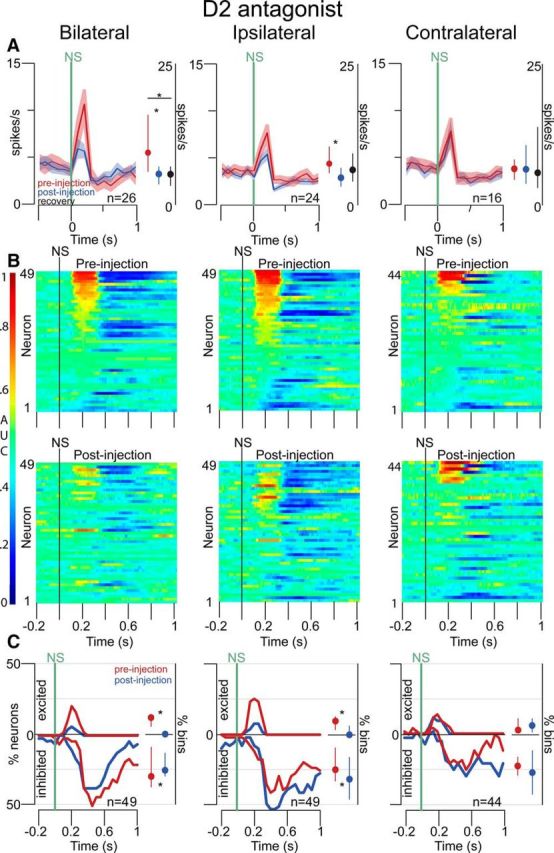
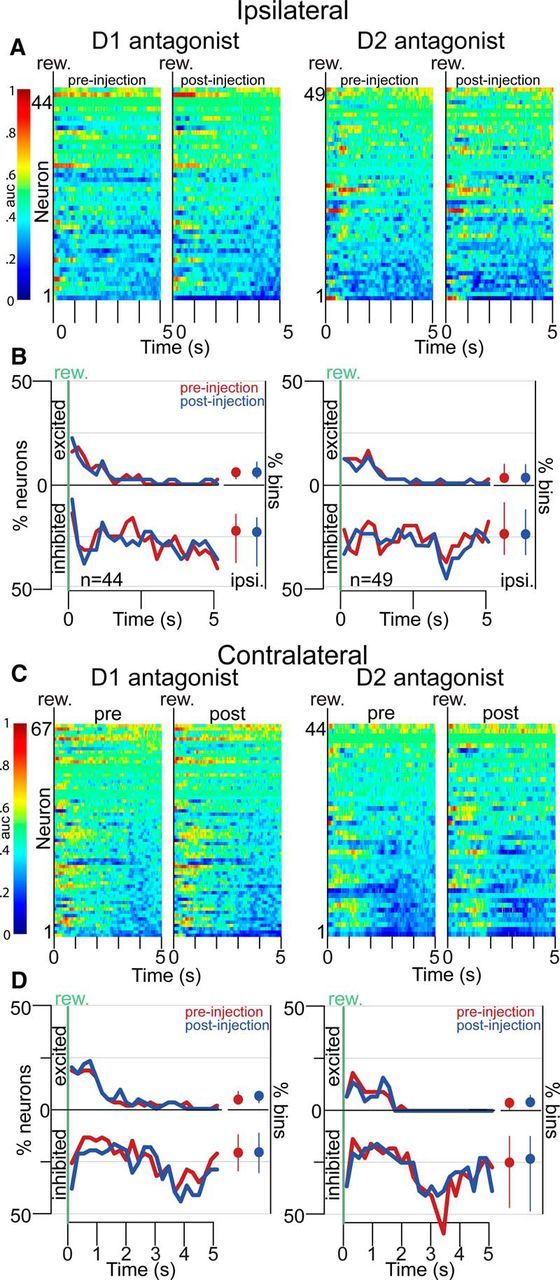
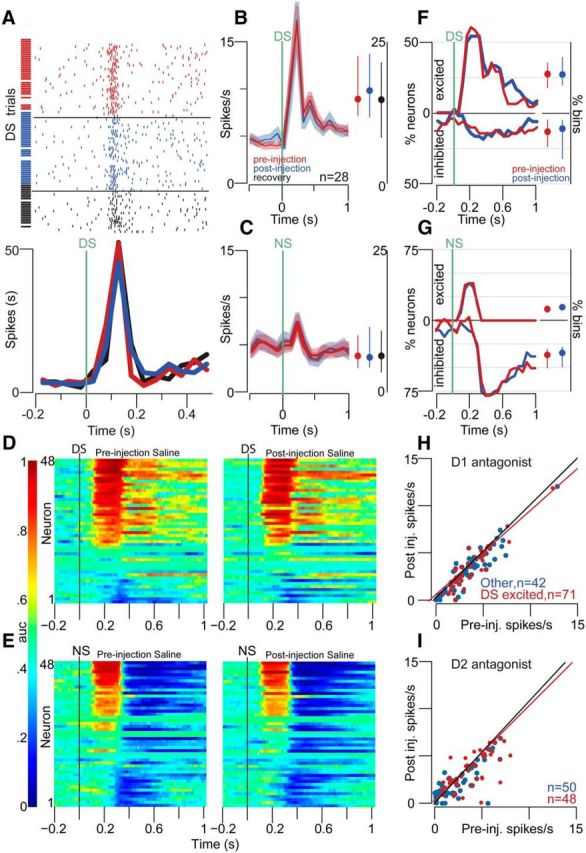
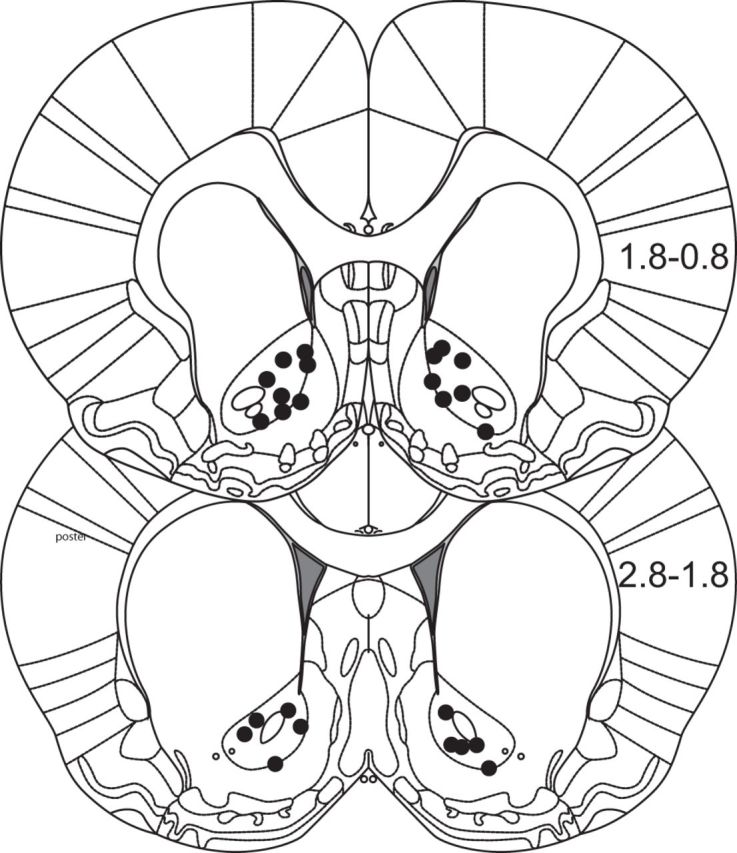
Approach to reward is a fundamental adaptive behavior, disruption of which is a core symptom of addiction and depression. Nucleus accumbens (NAc) dopamine is required for reward-predictive cues to activate vigorous reward seeking, but the underlying neural mechanism is unknown. Reward-predictive cues elicit both dopamine release in the NAc and excitations and inhibitions in NAc neurons. However, a direct link has not been established between dopamine receptor activation, NAc cue-evoked neuronal activity, and reward-seeking behavior. Here, we use a novel microelectrode array that enables simultaneous recording of neuronal firing and local dopamine receptor antagonist injection. We demonstrate that, in the NAc of rats performing a discriminative stimulus task for sucrose reward, blockade of either D1 or D2 receptors selectively attenuates excitation, but not inhibition, evoked by reward-predictive cues. Furthermore, we establish that this dopamine-dependent signal is necessary for reward-seeking behavior. These results demonstrate a neural mechanism by which NAc dopamine invigorates environmentally cued reward-seeking behavior.
Keywords: cue-excited neurons; discriminative stimulus; dopamine; nucleus accumbens; reward seeking.
Copyright © 2014 the authors 0270-6474/14/3414349-16$15.00/0.
Figures
References
-
- Brog JS, Salyapongse A, Deutch AY, Zahm DS. The patterns of afferent innervation of the core and shell in the “accumbens” part of the rat ventral striatum: immunohistochemical detection of retrogradely transported fluoro-gold. J Comp Neurol. 1993;338:255–278. doi: 10.1002/cne.903380209. - DOI - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources