

Isolating neural correlates of conscious perception from neural correlates of reporting one's perception
- PMID: 25339922
- PMCID: PMC4189413
- DOI: 10.3389/fpsyg.2014.01078
Isolating neural correlates of conscious perception from neural correlates of reporting one's perception
Abstract
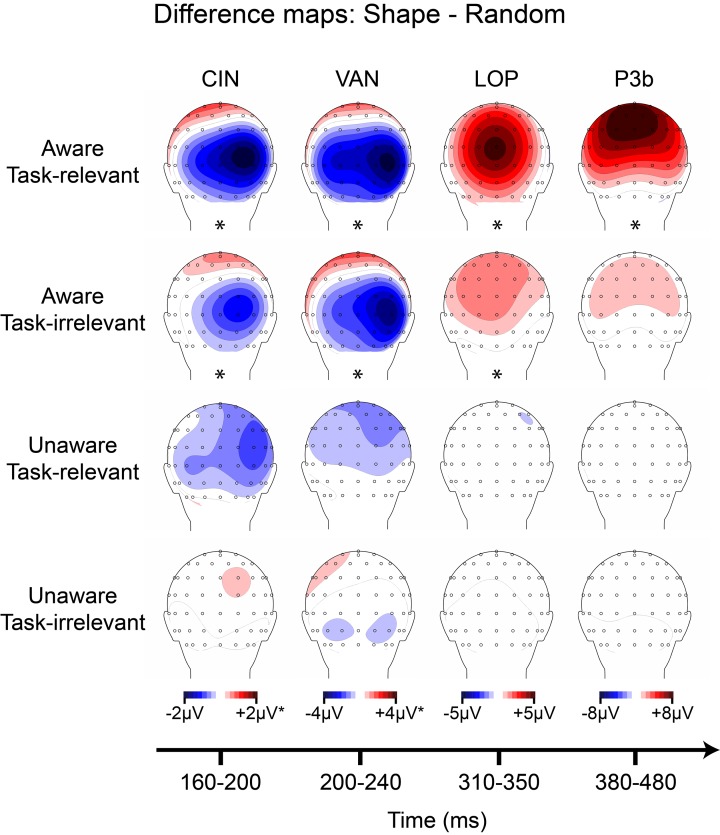
To isolate neural correlates of conscious perception (NCCs), a standard approach has been to contrast neural activity elicited by identical stimuli of which subjects are aware vs. unaware. Because conscious experience is private, determining whether a stimulus was consciously perceived requires subjective report: e.g., button-presses indicating detection, visibility ratings, verbal reports, etc. This reporting requirement introduces a methodological confound when attempting to isolate NCCs: The neural processes responsible for accessing and reporting one's percept are difficult to distinguish from those underlying the conscious percept itself. Here, we review recent attempts to circumvent this issue via a modified inattentional blindness paradigm (Pitts et al., 2012) and present new data from a backward masking experiment in which task-relevance and visual awareness were manipulated in a 2 × 2 crossed design. In agreement with our previous inattentional blindness results, stimuli that were consciously perceived yet not immediately accessed for report (aware, task-irrelevant condition) elicited a mid-latency posterior ERP negativity (~200-240 ms), while stimuli that were accessed for report (aware, task-relevant condition) elicited additional components including a robust P3b (~380-480 ms) subsequent to the mid-latency negativity. Overall, these results suggest that some of the NCCs identified in previous studies may be more closely linked with accessing and maintaining perceptual information for reporting purposes than with encoding the conscious percept itself. An open question is whether the remaining NCC candidate (the ERP negativity at 200-240 ms) reflects visual awareness or object-based attention.
Keywords: P3b; VAN; attention; awareness; masking; task-relevance.
Figures








References
-
- Baars B. J. (1989). A Cognitive Theory of Consciousness. Cambridge, MA: Cambridge University Press
LinkOut - more resources
Full Text Sources
Other Literature Sources