

The Drosophila Tis11 protein and its effects on mRNA expression in flies
- PMID: 25342740
- PMCID: PMC4271196
- DOI: 10.1074/jbc.M114.593491
The Drosophila Tis11 protein and its effects on mRNA expression in flies
Abstract
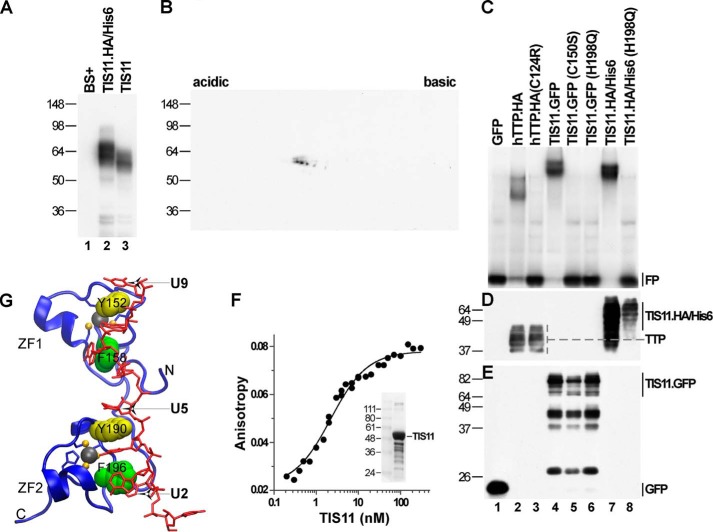
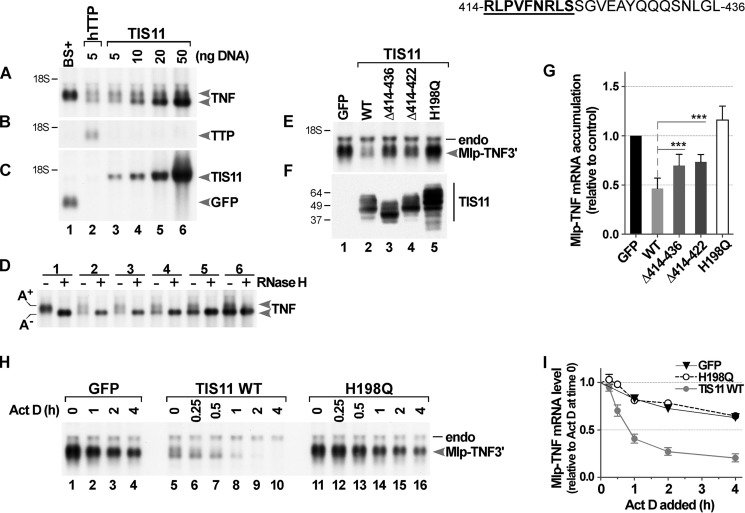
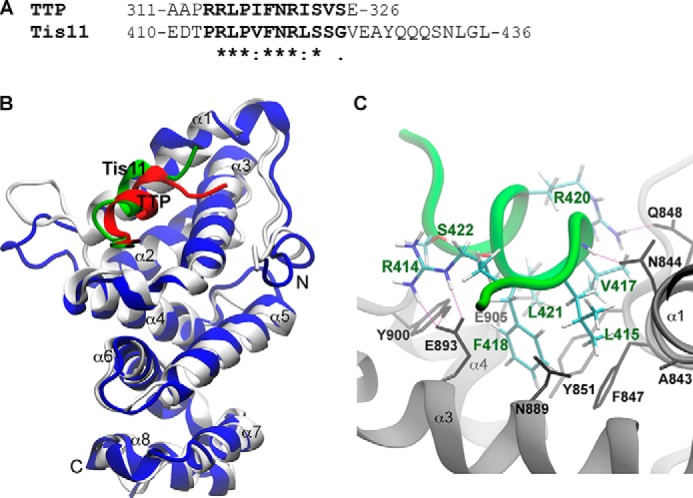
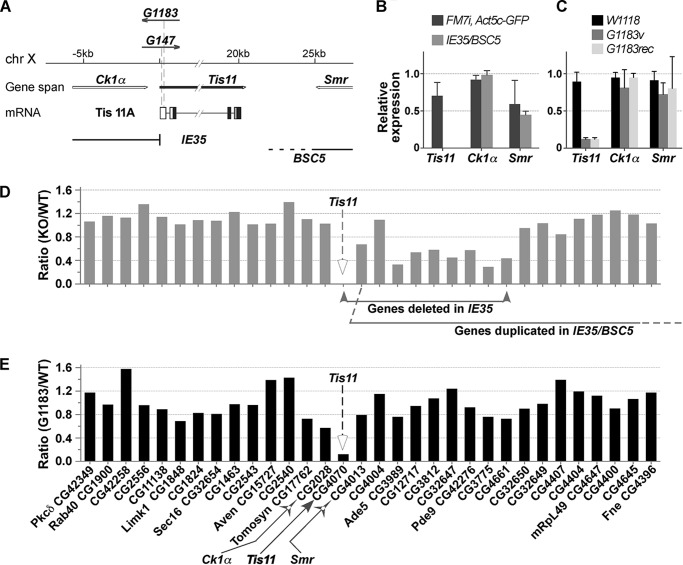
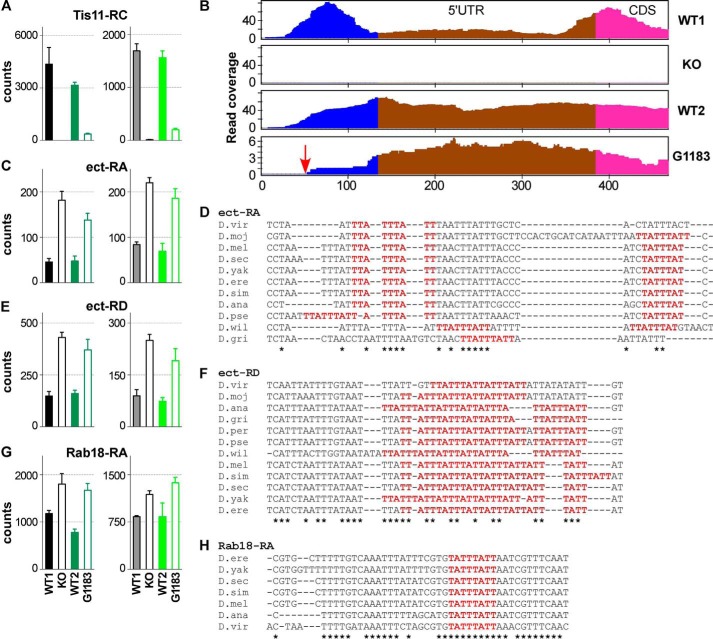
Members of the mammalian tristetraprolin family of CCCH tandem zinc finger proteins can bind to certain AU-rich elements (AREs) in mRNAs, leading to their deadenylation and destabilization. Mammals express three or four members of this family, but Drosophila melanogaster and other insects appear to contain a single gene, Tis11. We found that recombinant Drosophila Tis11 protein could bind to ARE-containing RNA oligonucleotides with low nanomolar affinity. Remarkably, co-expression in mammalian cells with "target" RNAs demonstrated that Tis11 could promote destabilization of ARE-containing mRNAs and that this was partially dependent on a conserved C-terminal sequence resembling the mammalian NOT1 binding domain. Drosophila Tis11 promoted both deadenylation and decay of a target transcript in this heterologous cell system. We used chromosome deletion/duplication and P element insertion to produce two types of Tis11 deficiency in adult flies, both of which were viable and fertile. To address the hypothesis that Tis11 deficiency would lead to the abnormal accumulation of potential target transcripts, we analyzed gene expression in adult flies by deep mRNA sequencing. We identified 69 transcripts from 56 genes that were significantly up-regulated more than 1.5-fold in both types of Tis11-deficient flies. Ten of the up-regulated transcripts encoded probable proteases, but many other functional classes of proteins were represented. Many of the up-regulated transcripts contained potential binding sites for tristetraprolin family member proteins that were conserved in other Drosophila species. Tis11 is thus an ARE-binding, mRNA-destabilizing protein that may play a role in post-transcriptional gene expression in Drosophila and other insects.
Keywords: AU-rich Elements; Deadenylation; Drosophila; Post-transcriptional Regulation; RNA-binding Protein; Zinc Finger; mRNA Decay.
© 2014 by The American Society for Biochemistry and Molecular Biology, Inc.
Figures

References
-
- Shaw G., Kamen R. (1986) A conserved AU sequence from the 3′ untranslated region of GM-CSF mRNA mediates selective mRNA degradation. Cell 46, 659–667 - PubMed
-
- Chen C. Y., Shyu A. B. (1995) AU-rich elements: characterization and importance in mRNA degradation. Trends Biochem. Sci. 20, 465–470 - PubMed
-
- Blackshear P. J. (2002) Tristetraprolin and other CCCH tandem zinc-finger proteins in the regulation of mRNA turnover. Biochem. Soc. Trans. 30, 945–952 - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
