

High thermodynamic stability of parametrically designed helical bundles
- PMID: 25342806
- PMCID: PMC4612401
- DOI: 10.1126/science.1257481
High thermodynamic stability of parametrically designed helical bundles
Abstract
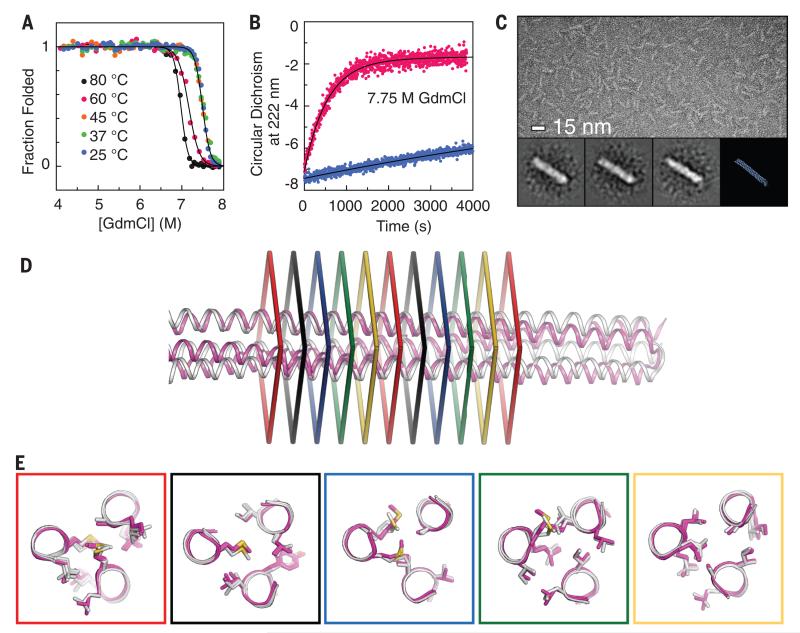
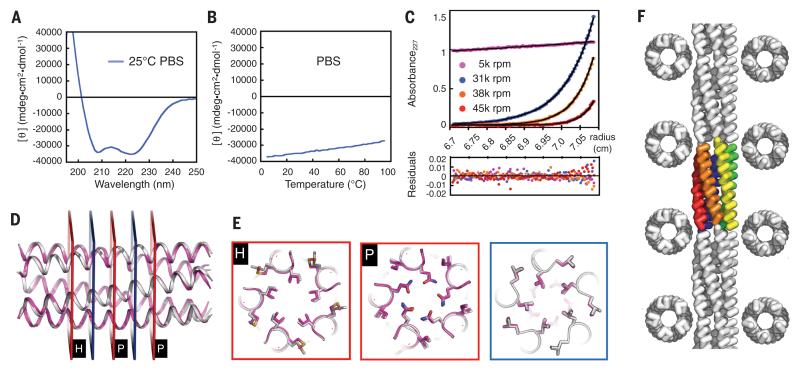
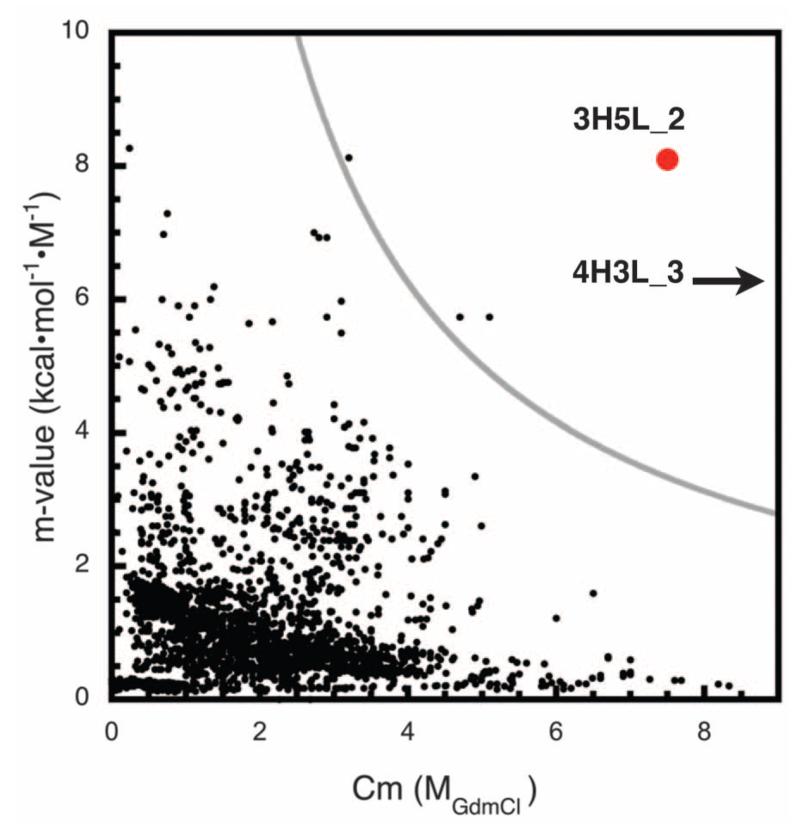
We describe a procedure for designing proteins with backbones produced by varying the parameters in the Crick coiled coil-generating equations. Combinatorial design calculations identify low-energy sequences for alternative helix supercoil arrangements, and the helices in the lowest-energy arrangements are connected by loop building. We design an antiparallel monomeric untwisted three-helix bundle with 80-residue helices, an antiparallel monomeric right-handed four-helix bundle, and a pentameric parallel left-handed five-helix bundle. The designed proteins are extremely stable (extrapolated ΔGfold > 60 kilocalories per mole), and their crystal structures are close to those of the design models with nearly identical core packing between the helices. The approach enables the custom design of hyperstable proteins with fine-tuned geometries for a wide range of applications.
Copyright © 2014, American Association for the Advancement of Science.
Figures

References
Publication types
MeSH terms
Associated data
- Actions
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
