

ERG oncoprotein inhibits ANXA2 expression and function in prostate cancer
- PMID: 25344575
- PMCID: PMC4336823
- DOI: 10.1158/1541-7786.MCR-14-0275-T
ERG oncoprotein inhibits ANXA2 expression and function in prostate cancer
Abstract
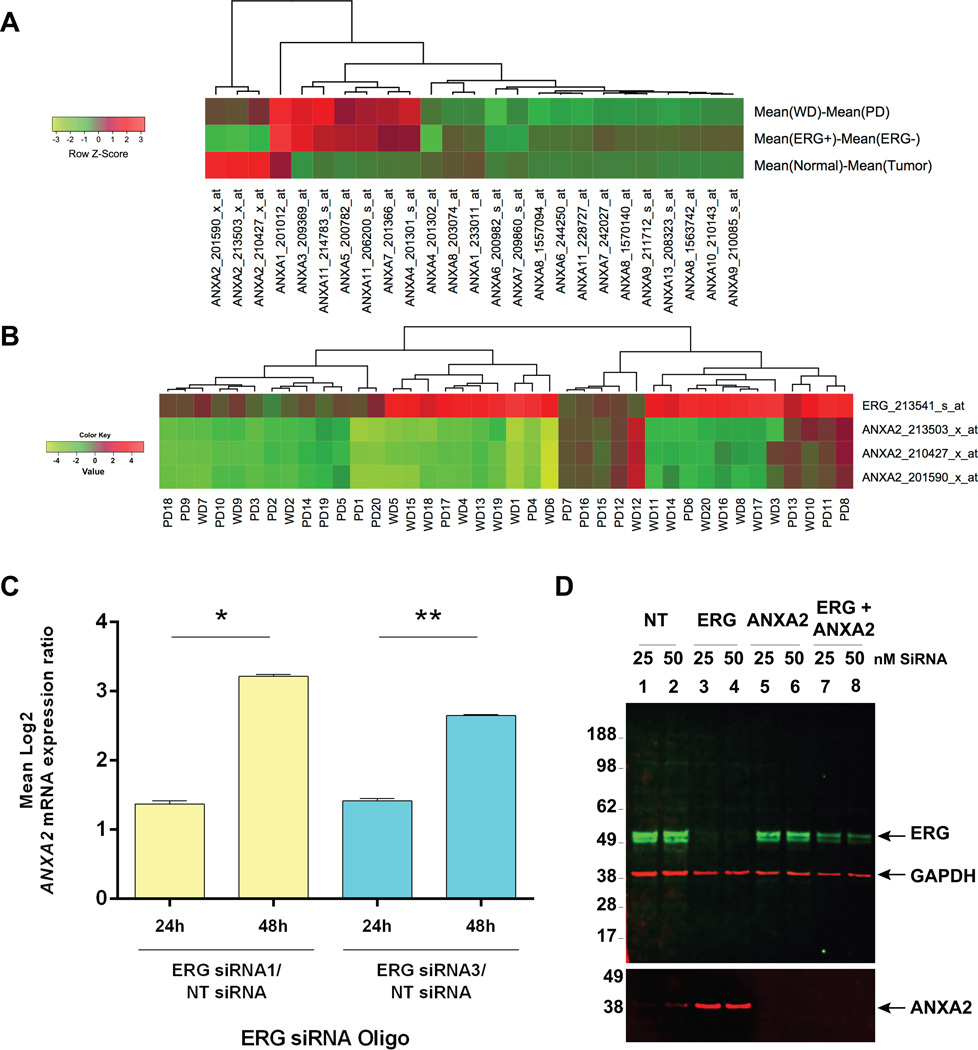
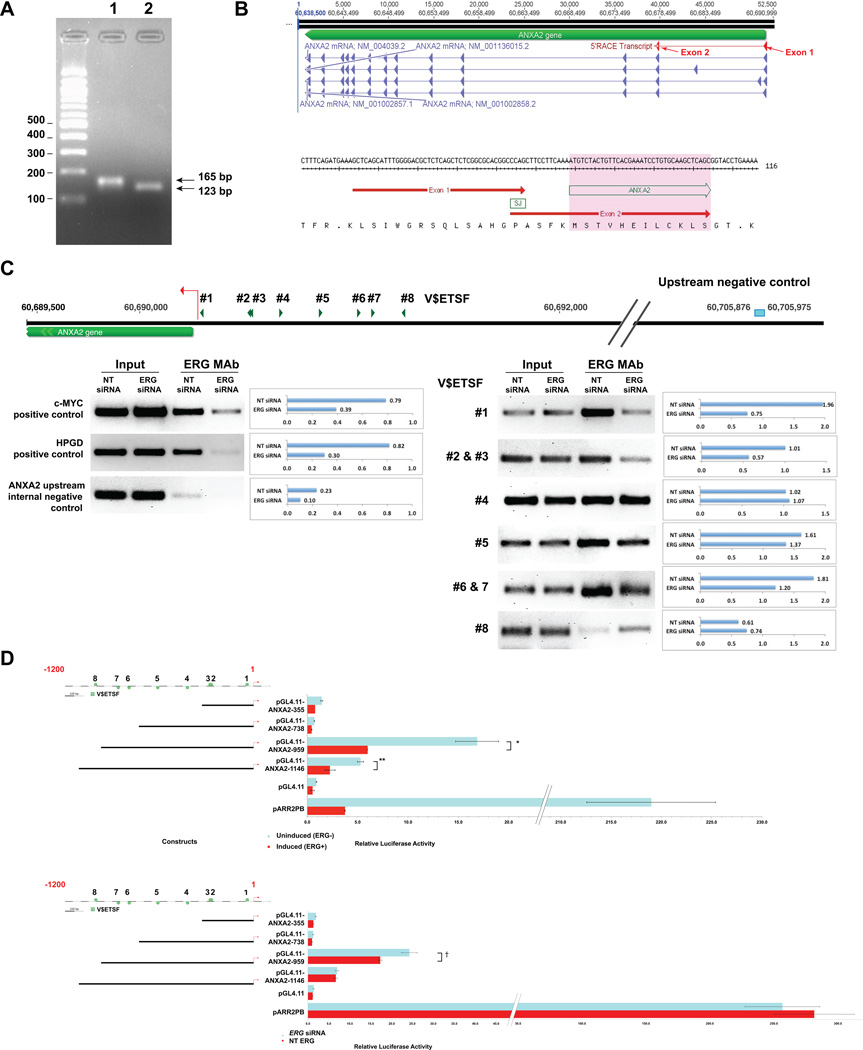
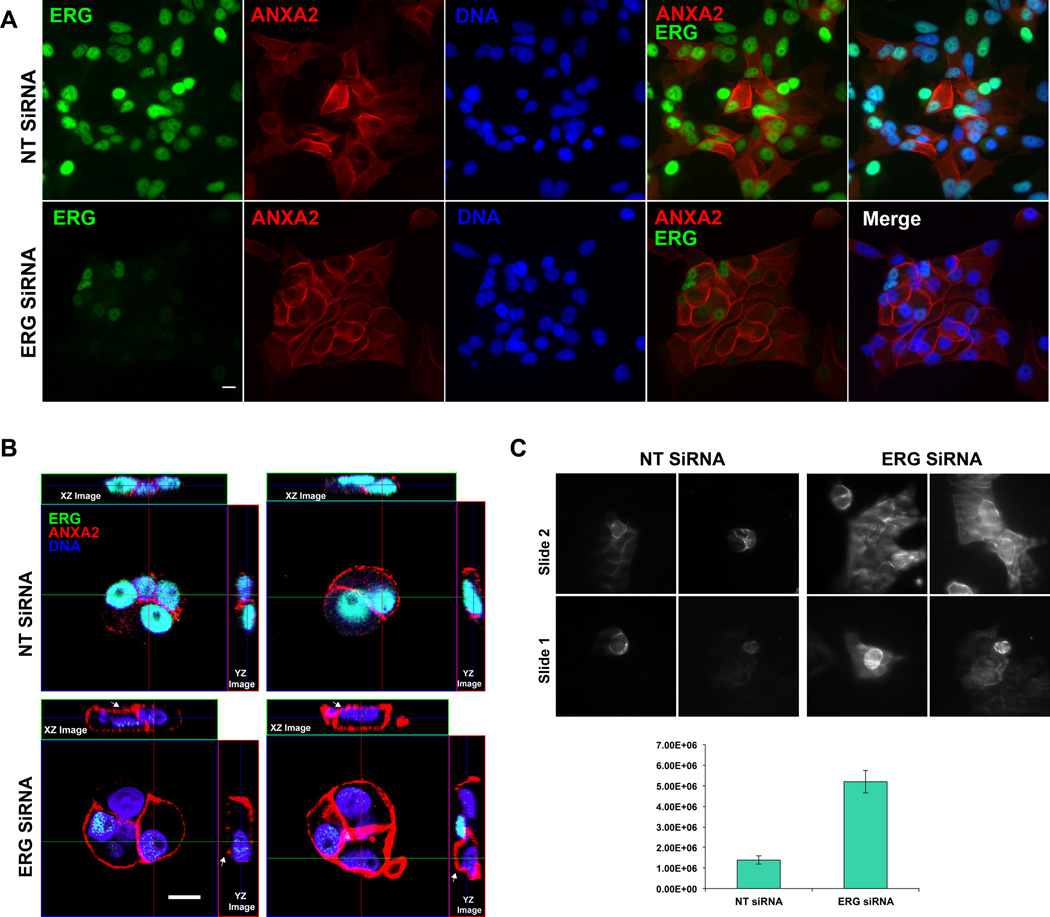
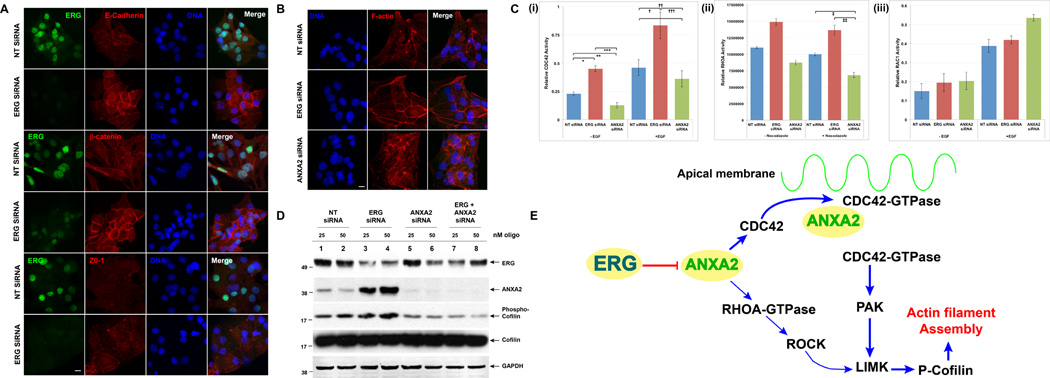
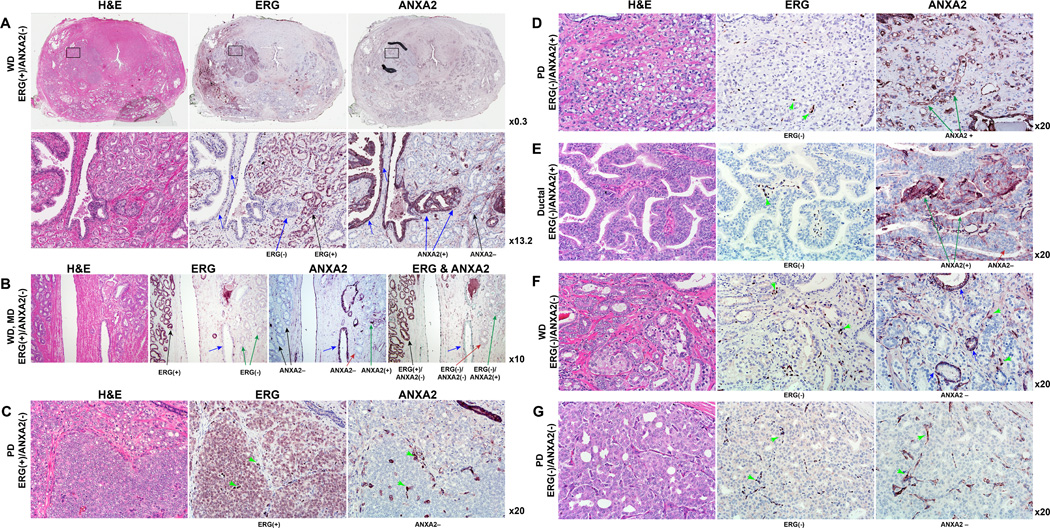
Overexpression of ERG in the prostate epithelium, due to chromosomal translocations, contributes to prostate tumorigenesis. Here, genomic analysis of ERG siRNA-treated prostate cells harboring the endogenous TMPRSS2-ERG fusion revealed an inverse relationship between ERG and Annexin A2 (ANXA2) expression at both the RNA and protein level. ANXA2, a Ca(2+)-dependent and phospholipid-binding protein, is involved in various cellular functions, including maintenance of epithelial cell polarity. Mechanistic studies defined the prostate-specific transcription start site of ANXA2 and showed that the recruitment of ERG to the ANXA2 promoter is required for transcriptional repression by ERG. Knockdown of ERG enhanced the apical localization of ANXA2, the bundling of actin filaments at cell-cell junctions and formation of a polarized epithelial phenotype. ERG overexpression disrupted ANXA2-mediated cell polarity and promoted epithelial-mesenchymal transition (EMT) by inhibiting CDC42 and RHOA, and by activating cofilin. Immunohistochemistry demonstrated a reciprocal relationship of ANXA2 and ERG expression in a large fraction of primary prostate cancer clinical specimens. ANXA2 was absent or markedly reduced in ERG(+) tumors, which were mostly well differentiated. ERG(-) tumors, meanwhile, expressed moderate to high levels of ANXA2, and were either poorly differentiated or displayed subsets of poorly differentiated cells. Taken together, the transcriptional repression of ANXA2 by ERG in prostate epithelial cells plays a critical role in abrogating differentiation, promoting EMT, and in the reciprocal correlation of ERG and ANXA2 expression observed in human prostate cancer.
Implications: ANXA2 is a new component of the ERG network with potential to enhance biologic stratification and therapeutic targeting of ERG-stratified prostate cancers.
©2014 American Association for Cancer Research.
Conflict of interest statement
The authors disclose no potential conflicts of interest.
Figures
Similar articles
-
Prognostic features of Annexin A2 expression in prostate cancer.Pathology. 2021 Feb;53(2):205-213. doi: 10.1016/j.pathol.2020.07.006. Epub 2020 Sep 20. Pathology. 2021. PMID: 32967771 Free PMC article.
-
ERG-SOX4 interaction promotes epithelial-mesenchymal transition in prostate cancer cells.Prostate. 2014 May;74(6):647-58. doi: 10.1002/pros.22783. Epub 2014 Jan 16. Prostate. 2014. PMID: 24435928
-
FZD4 as a mediator of ERG oncogene-induced WNT signaling and epithelial-to-mesenchymal transition in human prostate cancer cells.Cancer Res. 2010 Sep 1;70(17):6735-45. doi: 10.1158/0008-5472.CAN-10-0244. Epub 2010 Aug 16. Cancer Res. 2010. PMID: 20713528
-
ERG protein expression as a biomarker of prostate cancer.Biomark Med. 2013 Dec;7(6):851-65. doi: 10.2217/bmm.13.105. Biomark Med. 2013. PMID: 24266818 Review.
-
Protein phosphorylation and its role in the regulation of Annexin A2 function.Biochim Biophys Acta Gen Subj. 2017 Nov;1861(11 Pt A):2515-2529. doi: 10.1016/j.bbagen.2017.08.024. Epub 2017 Sep 1. Biochim Biophys Acta Gen Subj. 2017. PMID: 28867585 Review.
Cited by
-
Age and Tumor Differentiation-Associated Gene Expression Based Analysis of Non-Familial Prostate Cancers.Front Oncol. 2021 Jan 26;10:584280. doi: 10.3389/fonc.2020.584280. eCollection 2020. Front Oncol. 2021. PMID: 33575208 Free PMC article.
-
Role of actin-binding proteins in prostate cancer.Front Cell Dev Biol. 2024 Jul 11;12:1430386. doi: 10.3389/fcell.2024.1430386. eCollection 2024. Front Cell Dev Biol. 2024. PMID: 39055653 Free PMC article. Review.
-
Long non-coding RNA LUCAT1 promotes tumourigenesis by inhibiting ANXA2 phosphorylation in hepatocellular carcinoma.J Cell Mol Med. 2019 Mar;23(3):1873-1884. doi: 10.1111/jcmm.14088. Epub 2018 Dec 26. J Cell Mol Med. 2019. PMID: 30588744 Free PMC article.
-
Leucine zipper protein 2 serves as a prognostic biomarker for prostate cancer correlating with immune infiltration and epigenetic regulation.Heliyon. 2022 Sep 29;8(10):e10750. doi: 10.1016/j.heliyon.2022.e10750. eCollection 2022 Oct. Heliyon. 2022. PMID: 36217461 Free PMC article.
-
Differential CircRNA Expression Signatures May Serve as Potential Novel Biomarkers in Prostate Cancer.Front Cell Dev Biol. 2021 Feb 25;9:605686. doi: 10.3389/fcell.2021.605686. eCollection 2021. Front Cell Dev Biol. 2021. PMID: 33718350 Free PMC article.
References
-
- Tomlins SA, Rhodes DR, Perner S, Dhanasekaran SM, Mehra R, Sun XW, et al. Recurrent fusion of TMPRSS2 and ETS transcription factor genes in prostate cancer. Science. 2005;310:644–648. - PubMed
-
- Petrovics G, Liu A, Shaheduzzaman S, Furusato B, Sun C, Chen Y, et al. Frequent overexpression of ETS-related gene-1 (ERG1) in prostate cancer transcriptome. Oncogene. 2005;24:3847–3852. - PubMed
-
- Dobi A, Sreenath T, Srivastava S. Androgen-Dependent Oncogenic Activation of ETS Transcription Factors by Recurrent Gene Fusions in Prostate Cancer: Biological and Clinical Implications. In: Wang Z, editor. Androgen-Responsive Genes in Prostate Cancer. New York: Springer; 2013. pp. 307–328.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous
