

Multiple polyploidy events in the early radiation of nodulating and nonnodulating legumes
- PMID: 25349287
- PMCID: PMC4271530
- DOI: 10.1093/molbev/msu296
Multiple polyploidy events in the early radiation of nodulating and nonnodulating legumes
Abstract
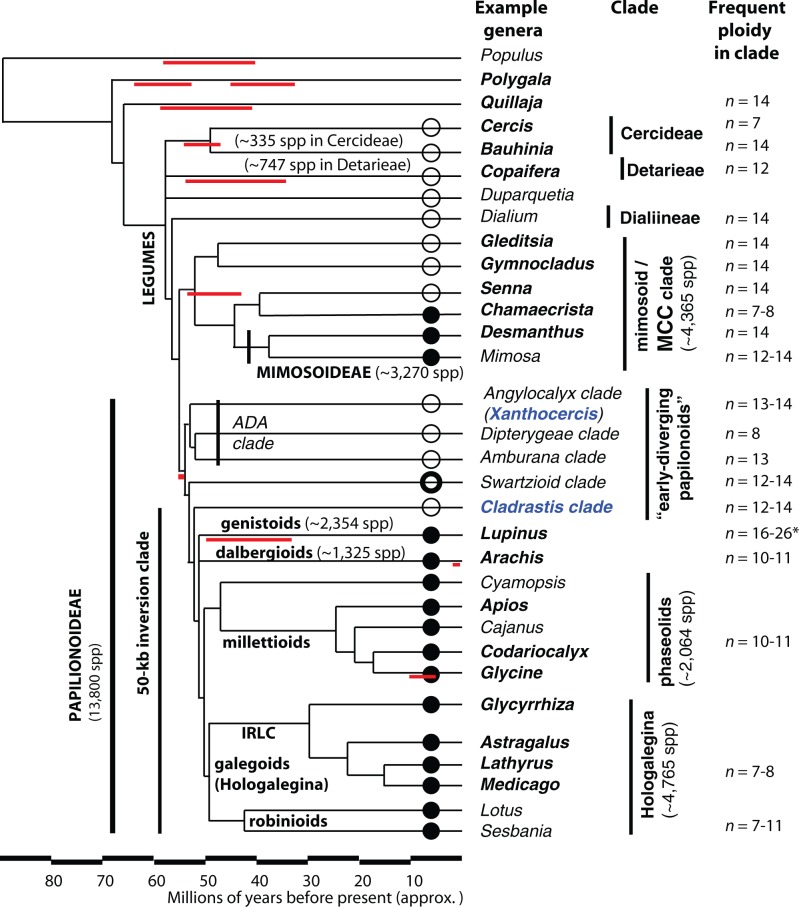
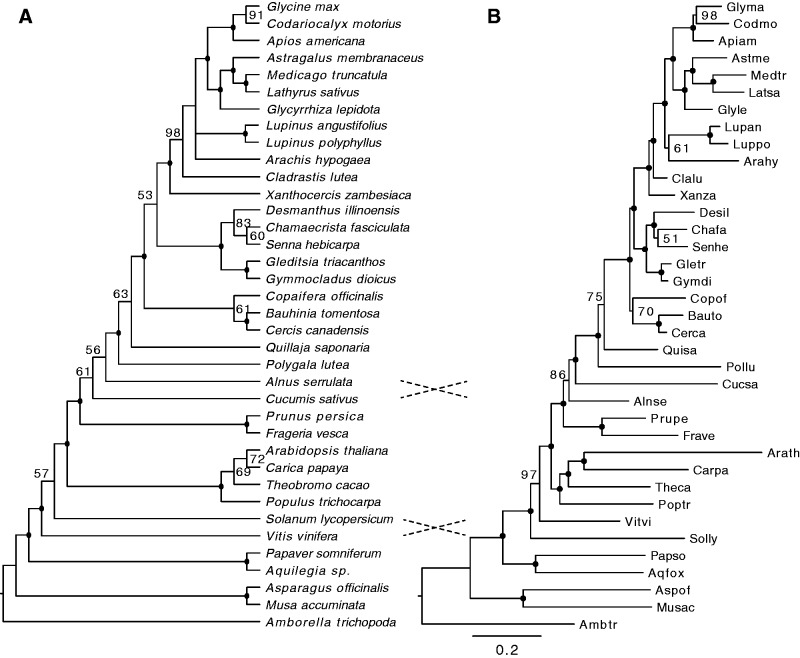
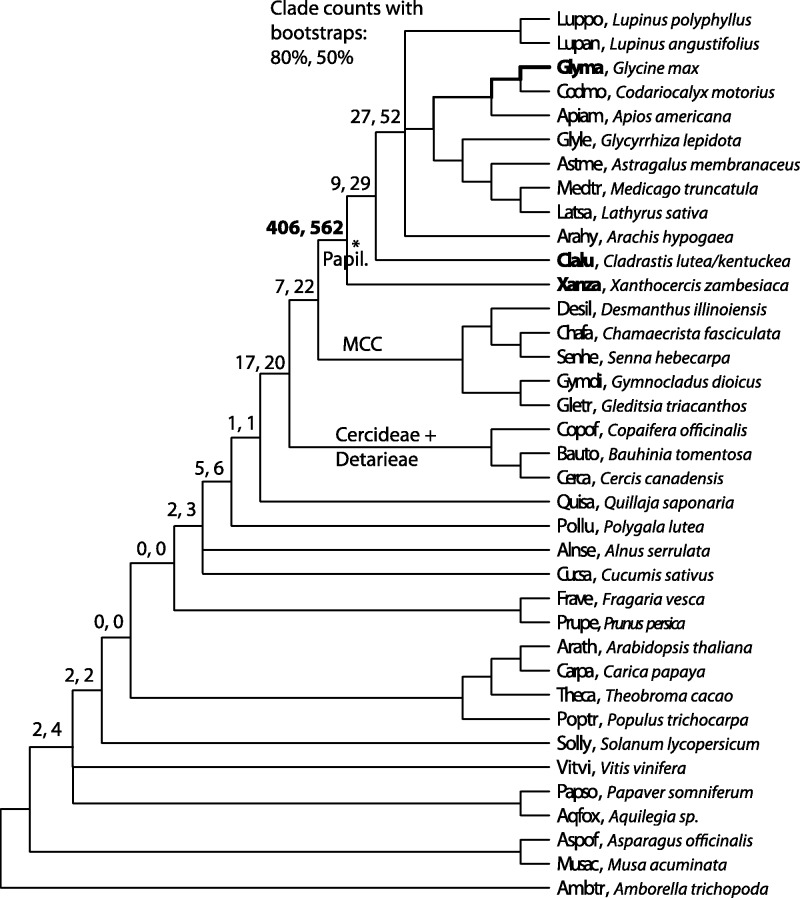
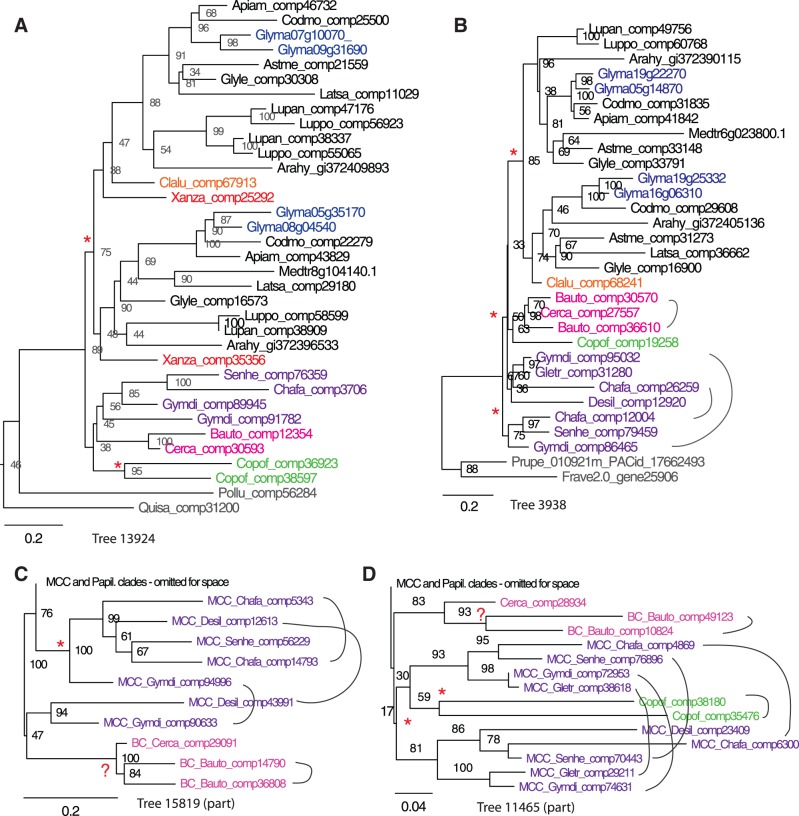
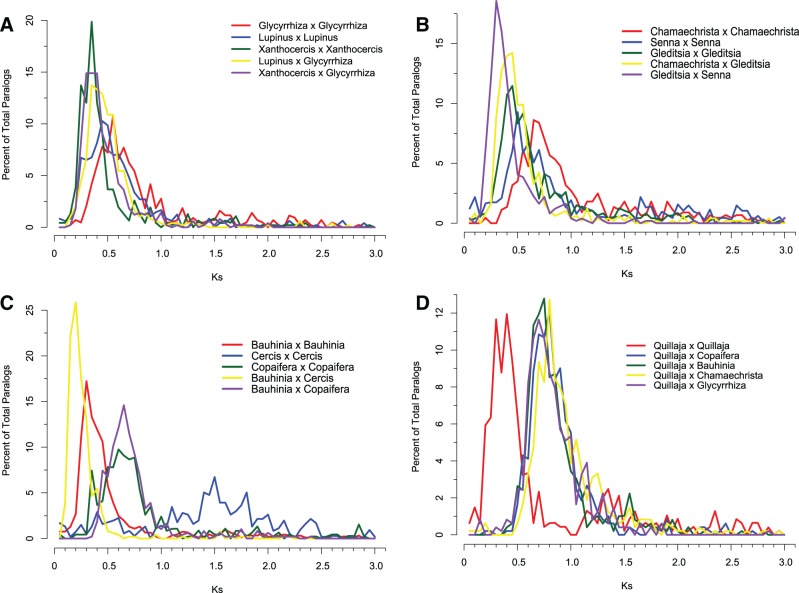
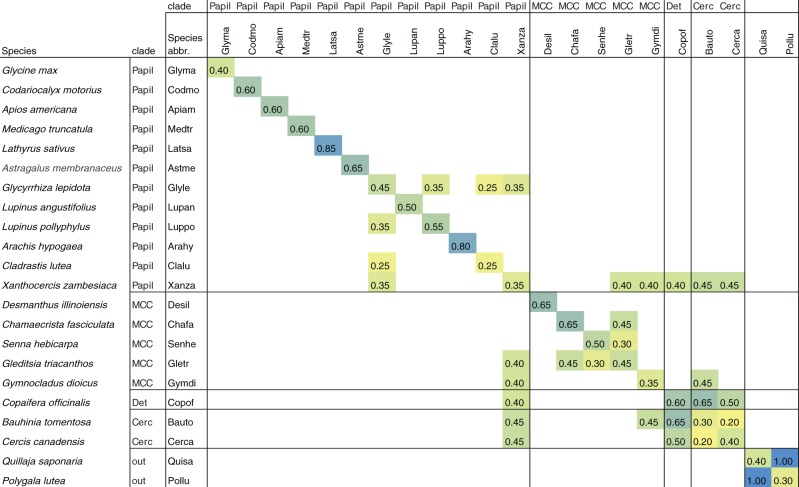
Unresolved questions about evolution of the large and diverse legume family include the timing of polyploidy (whole-genome duplication; WGDs) relative to the origin of the major lineages within the Fabaceae and to the origin of symbiotic nitrogen fixation. Previous work has established that a WGD affects most lineages in the Papilionoideae and occurred sometime after the divergence of the papilionoid and mimosoid clades, but the exact timing has been unknown. The history of WGD has also not been established for legume lineages outside the Papilionoideae. We investigated the presence and timing of WGDs in the legumes by querying thousands of phylogenetic trees constructed from transcriptome and genome data from 20 diverse legumes and 17 outgroup species. The timing of duplications in the gene trees indicates that the papilionoid WGD occurred in the common ancestor of all papilionoids. The earliest diverging lineages of the Papilionoideae include both nodulating taxa, such as the genistoids (e.g., lupin), dalbergioids (e.g., peanut), phaseoloids (e.g., beans), and galegoids (=Hologalegina, e.g., clovers), and clades with nonnodulating taxa including Xanthocercis and Cladrastis (evaluated in this study). We also found evidence for several independent WGDs near the base of other major legume lineages, including the Mimosoideae-Cassiinae-Caesalpinieae (MCC), Detarieae, and Cercideae clades. Nodulation is found in the MCC and papilionoid clades, both of which experienced ancestral WGDs. However, there are numerous nonnodulating lineages in both clades, making it unclear whether the phylogenetic distribution of nodulation is due to independent gains or a single origin followed by multiple losses.
Keywords: Mimosoideae; Papilionoideae; legume; nodulation; polyploidy; symbiotic nitrogen fixation.
Published by Oxford University Press on behalf of the Society for Molecular Biology and Evolution 2014. This work is written by US Government employees and is in the public domain in the US.
Figures
References
-
- Amborella Genome Project. The Amborella genome and the evolution of flowering plants. Science. 2013;342:1241089. - PubMed
-
- Ancestral Angiosperm Genome Project. 2013. [Internet]. Available: http://ancangio.uga.edu.
-
- Bell CD, Soltis DE, Soltis PS. The age and diversification of the angiosperms re-revisited. Am J Bot. 2010;97:1296–1303. - PubMed
-
- Bennett MD, Leitch IJ. 2012. Plant DNA C-values database (release 6.0, December 2012). [cited 2013 Dec]. Available from: http://www.kew.org/cvalues.
-
- Bertioli DJ, Moretzsohn MC, Madsen LH, Sandal N, Leal-Bertioli SC, Guimaraes PM, Hougaard BK, Fredslund J, Schauser L, Nielsen AM, et al. An analysis of synteny of Arachis with Lotus and Medicago sheds new light on the structure, stability and evolution of legume genomes. BMC Genomics. 2009;10:45. - PMC - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
