

Contraction-stimulated glucose transport in muscle is controlled by AMPK and mechanical stress but not sarcoplasmatic reticulum Ca(2+) release
- PMID: 25353002
- PMCID: PMC4209358
- DOI: 10.1016/j.molmet.2014.07.005
Contraction-stimulated glucose transport in muscle is controlled by AMPK and mechanical stress but not sarcoplasmatic reticulum Ca(2+) release
Abstract
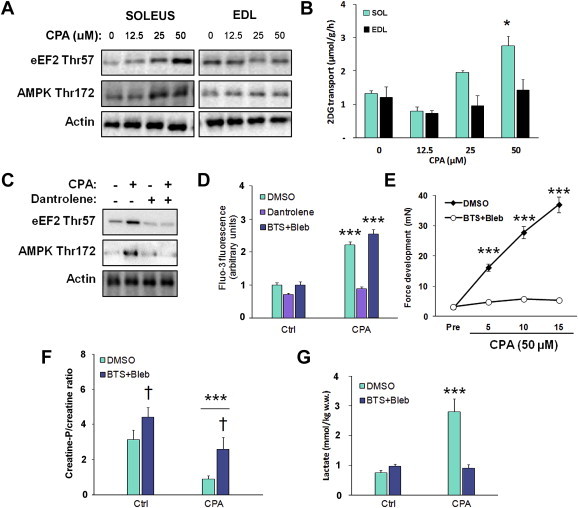
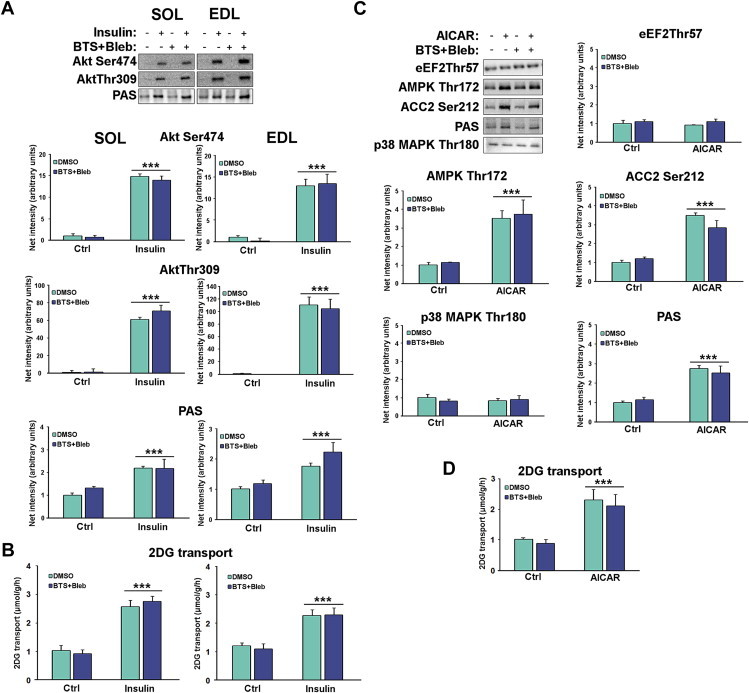
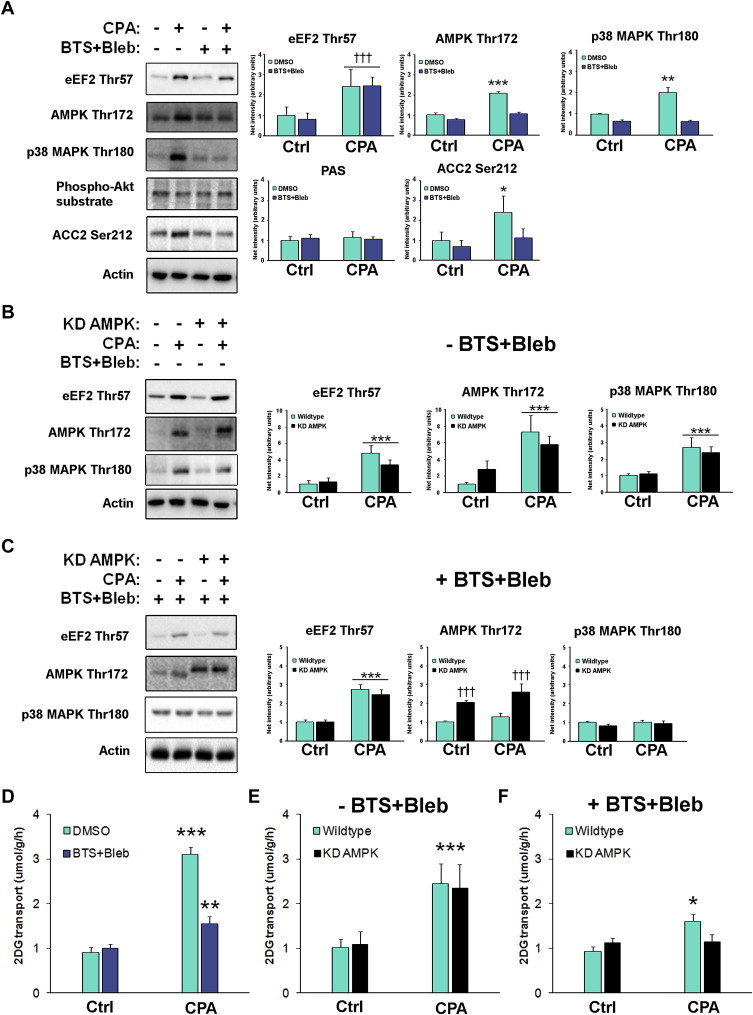
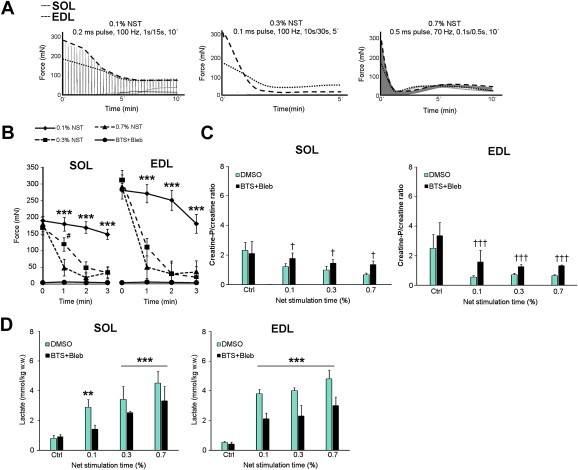
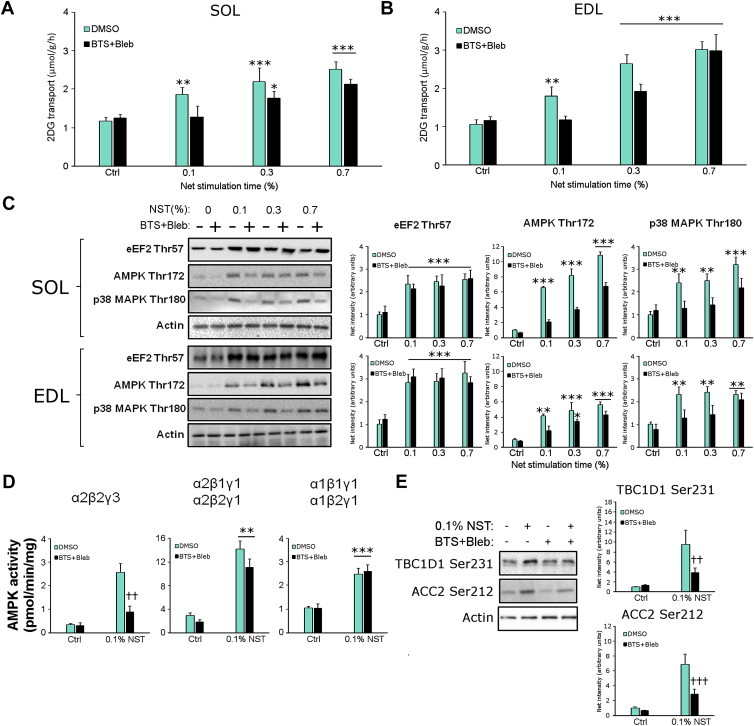
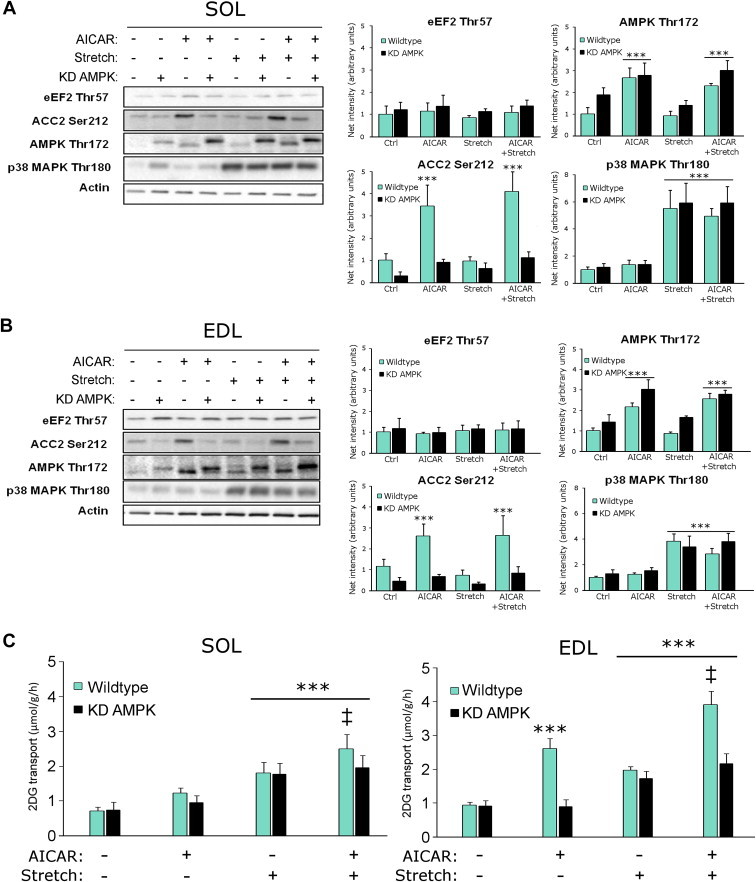
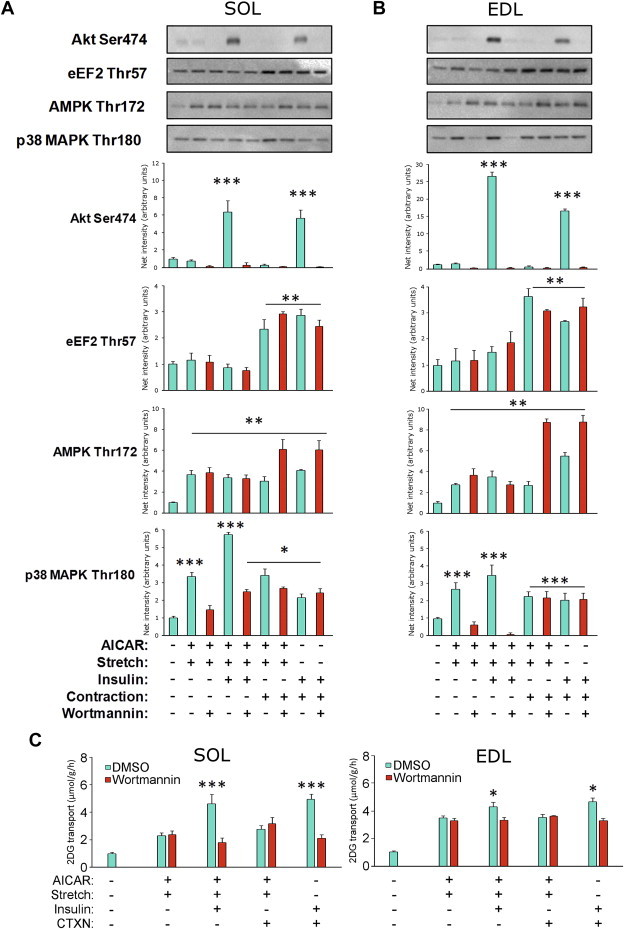
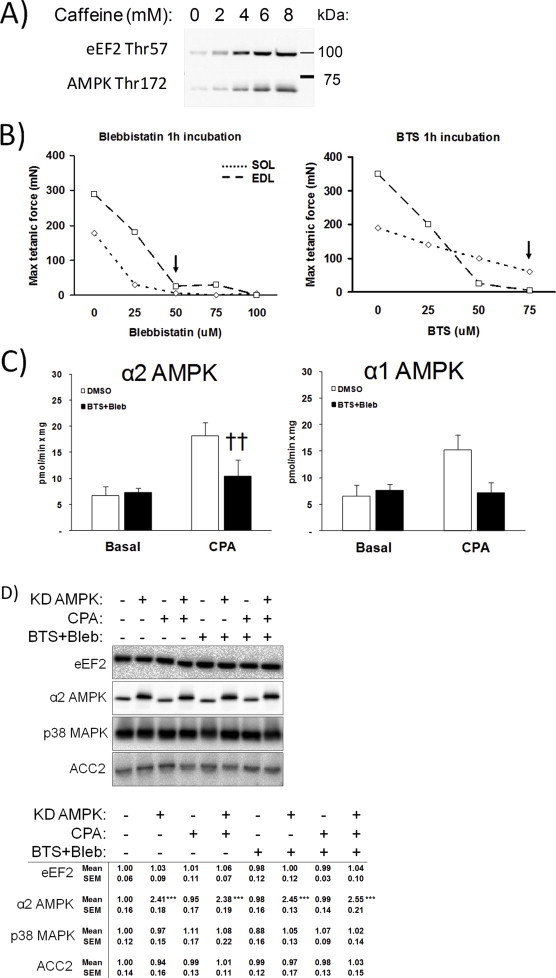
Understanding how muscle contraction orchestrates insulin-independent muscle glucose transport may enable development of hyperglycemia-treating drugs. The prevailing concept implicates Ca(2+) as a key feed forward regulator of glucose transport with secondary fine-tuning by metabolic feedback signals through proteins such as AMPK. Here, we demonstrate in incubated mouse muscle that Ca(2+) release is neither sufficient nor strictly necessary to increase glucose transport. Rather, the glucose transport response is associated with metabolic feedback signals through AMPK, and mechanical stress-activated signals. Furthermore, artificial stimulation of AMPK combined with passive stretch of muscle is additive and sufficient to elicit the full contraction glucose transport response. These results suggest that ATP-turnover and mechanical stress feedback are sufficient to fully increase glucose transport during muscle contraction, and call for a major reconsideration of the established Ca(2+) centric paradigm.
Keywords: AMPK; Ca2+; Exercise; Skeletal muscle; Stretch.
Figures
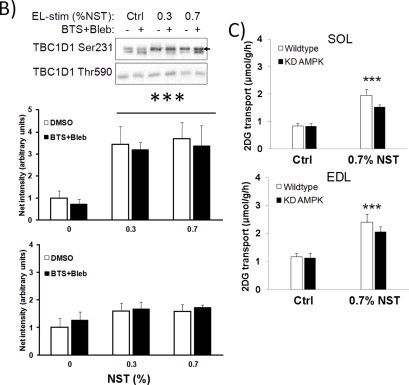
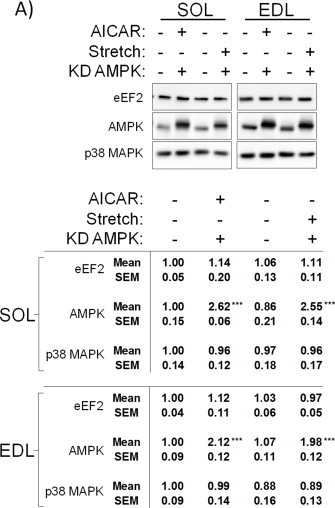
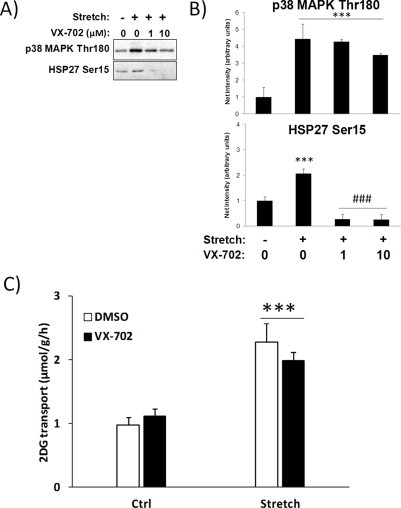
References
-
- James D.E., Kraegen E.W., Chisholm D.J. Muscle glucose metabolism in exercising rats: comparison with insulin stimulation. American Journal of Physiology. 1985;248:E575–E580. - PubMed
-
- Goodyear L.J., Kahn B.B. Exercise, glucose transport, and insulin sensitivity. Annual Review of Medicine. 1998;49:235–261. - PubMed
-
- Richter E.A., Hargreaves M. Exercise, GLUT4, and skeletal muscle glucose uptake. Physiological Reviews. 2013;93:993–1017. - PubMed
-
- Mu J., Brozinick J.T., Jr., Valladares O., Bucan M., Birnbaum M.J. A role for AMP-activated protein kinase in contraction- and hypoxia-regulated glucose transport in skeletal muscle. Molecular Cell. 2001;7:1085–1094. - PubMed
-
- Ihlemann J., Ploug T., Hellsten Y., Galbo H. Effect of tension on contraction-induced glucose transport in rat skeletal muscle. American Journal of Physiology. 1999;277:E208–E214. - PubMed
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
