

Mechanisms controlling neuromuscular junction stability
- PMID: 25359233
- PMCID: PMC11113273
- DOI: 10.1007/s00018-014-1768-z
Mechanisms controlling neuromuscular junction stability
Abstract
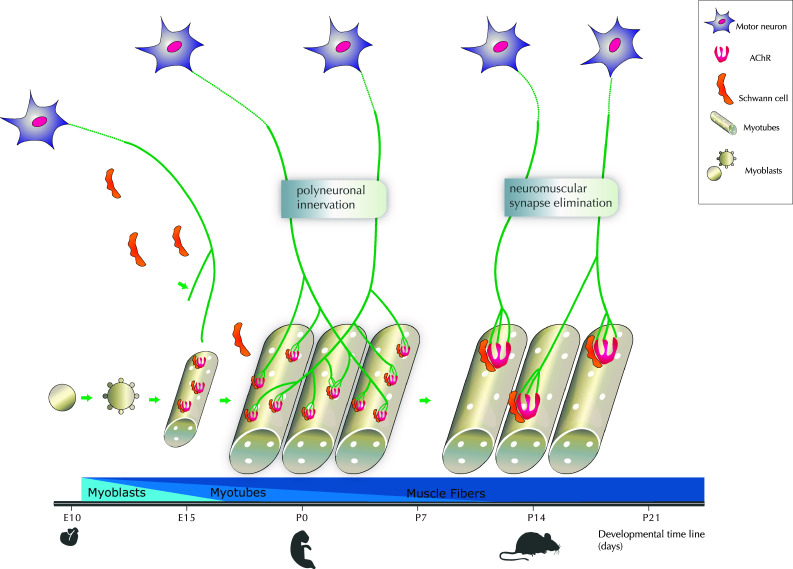
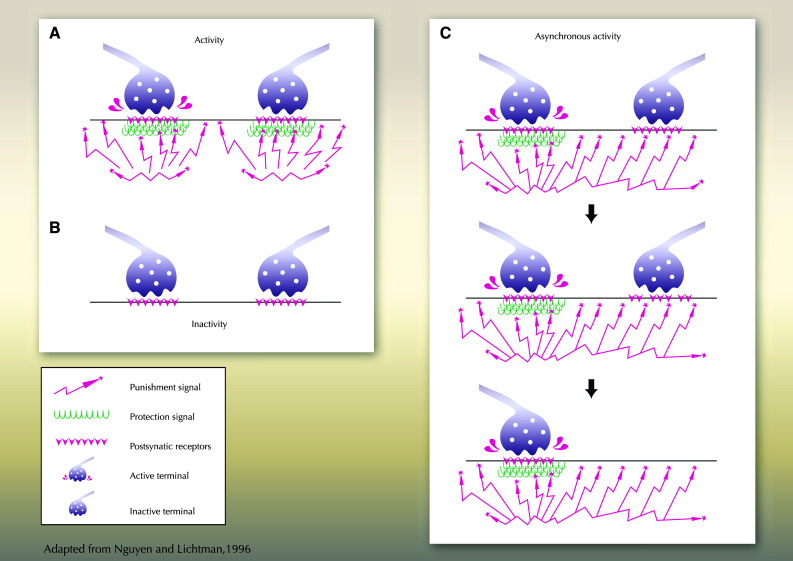
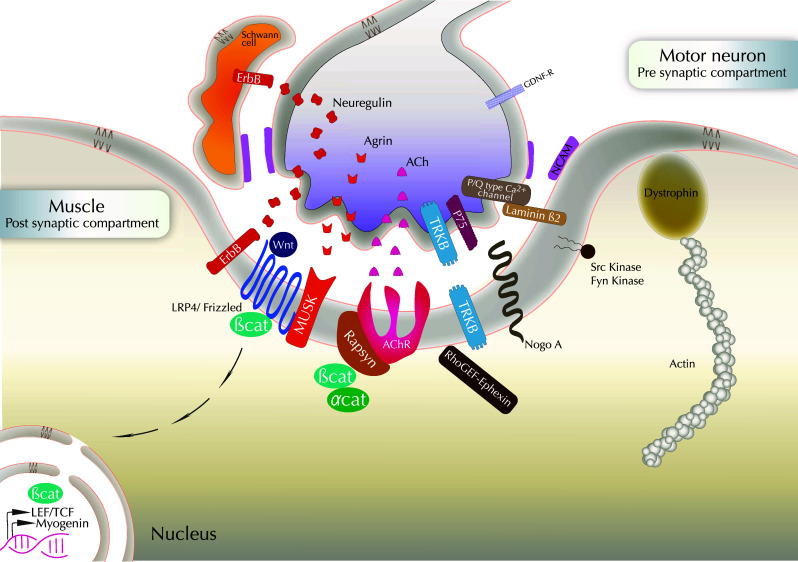
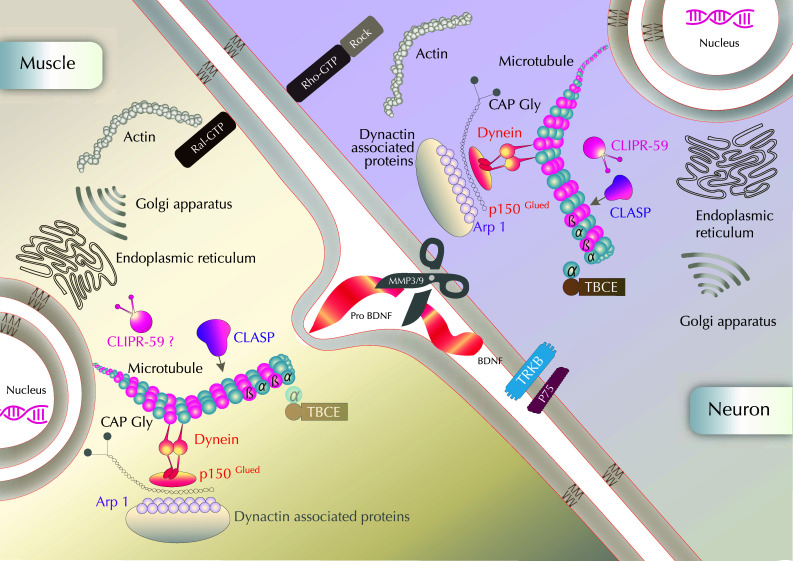
The neuromuscular junction (NMJ) is the synaptic connection between motor neurons and muscle fibers. It is involved in crucial processes such as body movements and breathing. Its proper development requires the guidance of motor axons toward their specific targets, the development of multi-innervated myofibers, and a selective synapse stabilization. It first consists of the removal of excessive motor axons on myofibers, going from multi-innervation to a single innervation of each myofiber. Whereas guidance cues of motor axons toward their specific muscular targets are well characterized, only few molecular and cellular cues have been reported as clues for selecting and stabilizing specific neuromuscular junctions. We will first provide a brief summary on NMJ development. We will then review molecular cues that are involved in NMJ stabilization, in both pre- and post-synaptic compartments, considering motor neurons and Schwann cells on the one hand, and muscle on the other hand. We will provide links with pathologies and highlight advances that can be brought both by basic research on NMJ development and clinical data resulting from the analyses of neurodegeneration of synaptic connections to obtain a better understanding of this process. The goal of this review is to highlight the findings toward understanding the roles of poly- or single-innervations and the underlying mechanisms of NMJ stabilization.
Conflict of interest statement
The authors declare no competing financial interests.
Figures
References
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
