

The role of the strictly conserved positively charged residue differs among the Gram-positive, Gram-negative, and chloroplast YidC homologs
- PMID: 25359772
- PMCID: PMC4271247
- DOI: 10.1074/jbc.M114.595082
The role of the strictly conserved positively charged residue differs among the Gram-positive, Gram-negative, and chloroplast YidC homologs
Abstract
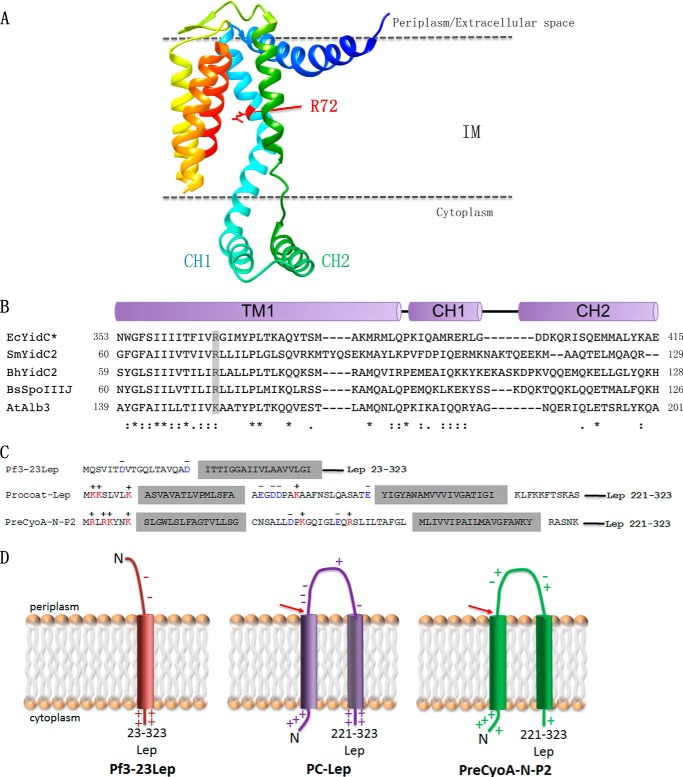
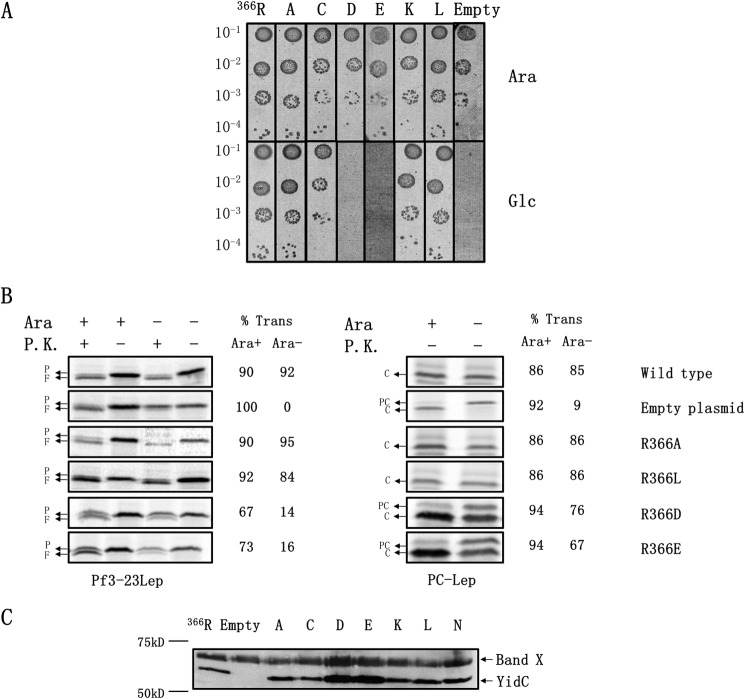
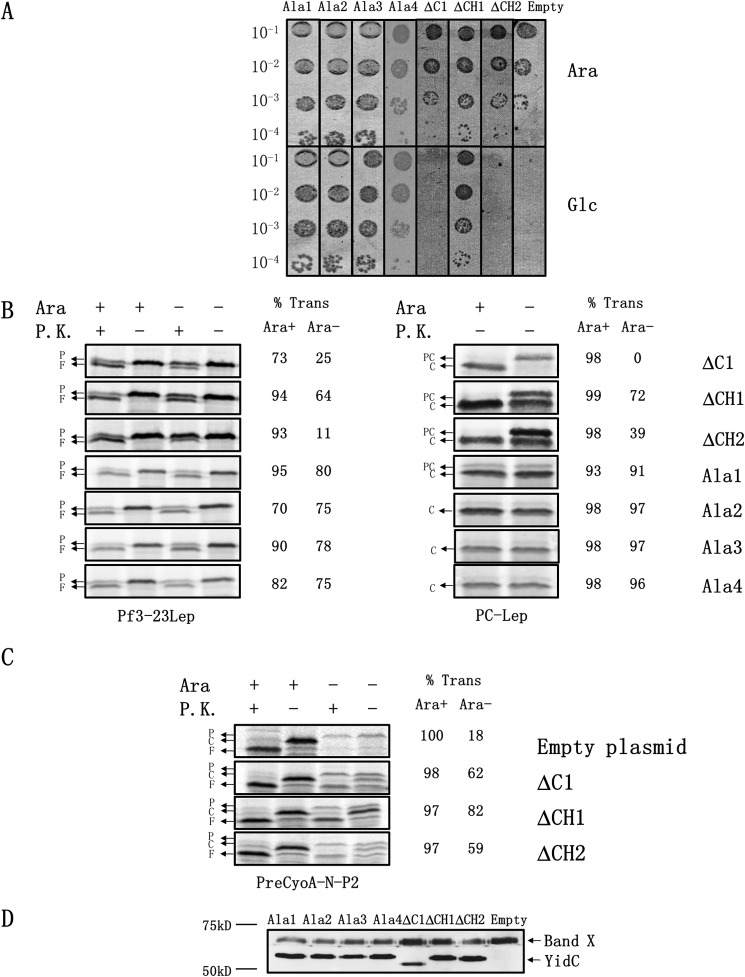
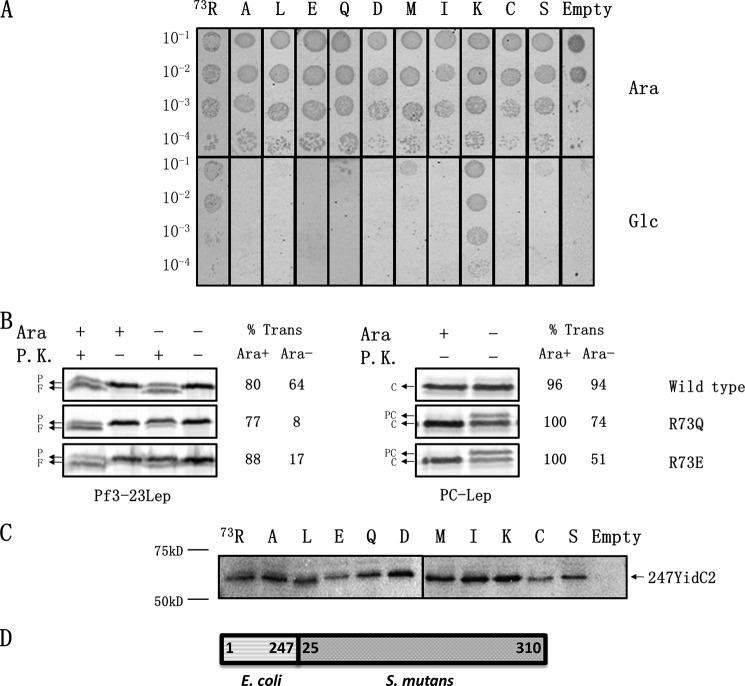
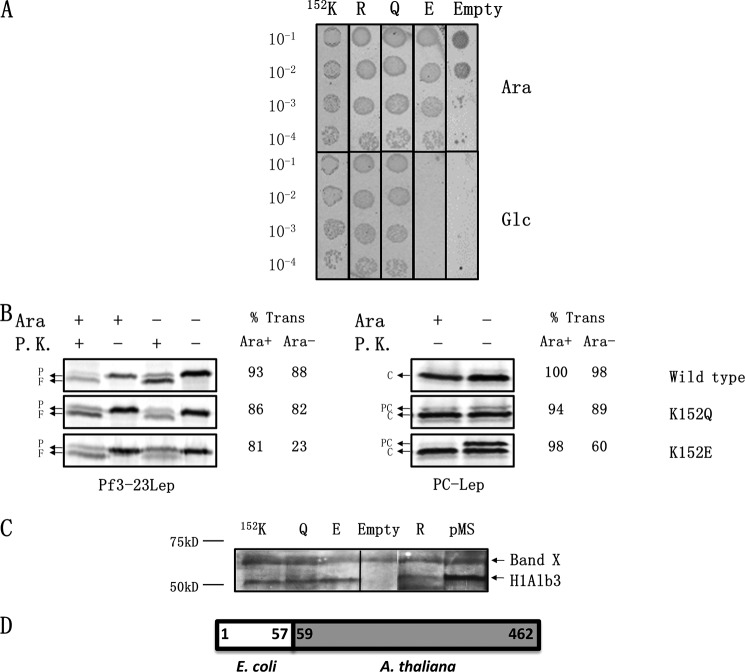
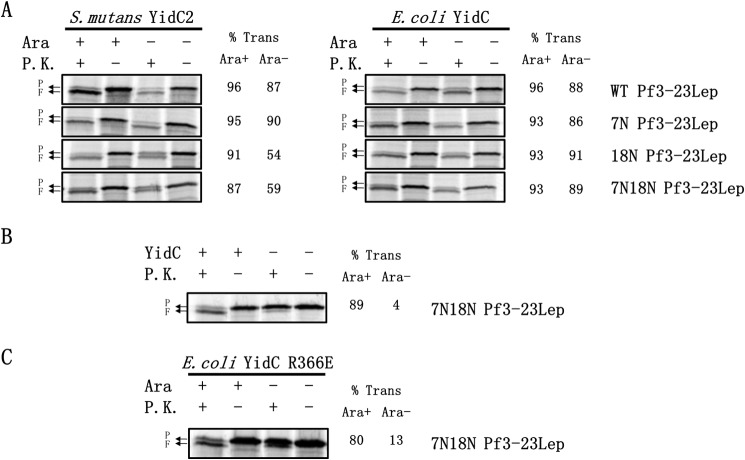
Recently, the structure of YidC2 from Bacillus halodurans revealed that the conserved positively charged residue within transmembrane segment one (at position 72) is located in a hydrophilic groove that is embedded in the inner leaflet of the lipid bilayer. The arginine residue was essential for the Bacillus subtilis SpoIIIJ (YidC1) to insert MifM and to complement a SpoIIIJ mutant strain. Here, we investigated the importance of the conserved positively charged residue for the function of the Escherichia coli YidC, Streptococcus mutans YidC2, and the chloroplast Arabidopsis thaliana Alb3. Like the Gram-positive B. subtilis SpoIIIJ, the conserved arginine was required for functioning of the Gram-positive S. mutans YidC2 and was necessary to complement the E. coli YidC depletion strain and to promote insertion of a YidC-dependent membrane protein synthesized with one but not two hydrophobic segments. In contrast, the conserved positively charged residue was not required for the E. coli YidC or the A. thaliana Alb3 to functionally complement the E. coli YidC depletion strain or to promote insertion of YidC-dependent membrane proteins. Our results also show that the C-terminal half of the helical hairpin structure in cytoplasmic loop C1 is important for the activity of YidC because various deletions in the region either eliminate or impair YidC function. The results here underscore the importance of the cytoplasmic hairpin region for YidC and show that the arginine is critical for the tested Gram-positive YidC homolog but is not essential for the tested Gram-negative and chloroplast YidC homologs.
Keywords: Alb3; Membrane Biogenesis; Membrane Enzyme; Membrane Function; Membrane Insertion; Membrane Protein; Membrane Structure; Structure-Function; YidC; YidC Mechanism.
© 2014 by The American Society for Biochemistry and Molecular Biology, Inc.
Figures
References
-
- Wang P., Dalbey R. E. (2011) Inserting membrane proteins: the YidC/Oxa1/Alb3 machinery in bacteria, mitochondria, and chloroplasts. Biochim. Biophys. Acta 1808, 866–875 - PubMed
-
- Kiefer D., Kuhn A. (2007) YidC as an essential and multifunctional component in membrane protein assembly. Int. Rev. Cytol. 259, 113–138 - PubMed
-
- Dalbey R. E., Kuhn A., Zhu L., Kiefer D. (2014) The membrane insertase YidC. Biochim. Biophys. Acta 1843, 1489–1496 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
