

Pannexin 1 regulates bidirectional hippocampal synaptic plasticity in adult mice
- PMID: 25360084
- PMCID: PMC4197765
- DOI: 10.3389/fncel.2014.00326
Pannexin 1 regulates bidirectional hippocampal synaptic plasticity in adult mice
Abstract
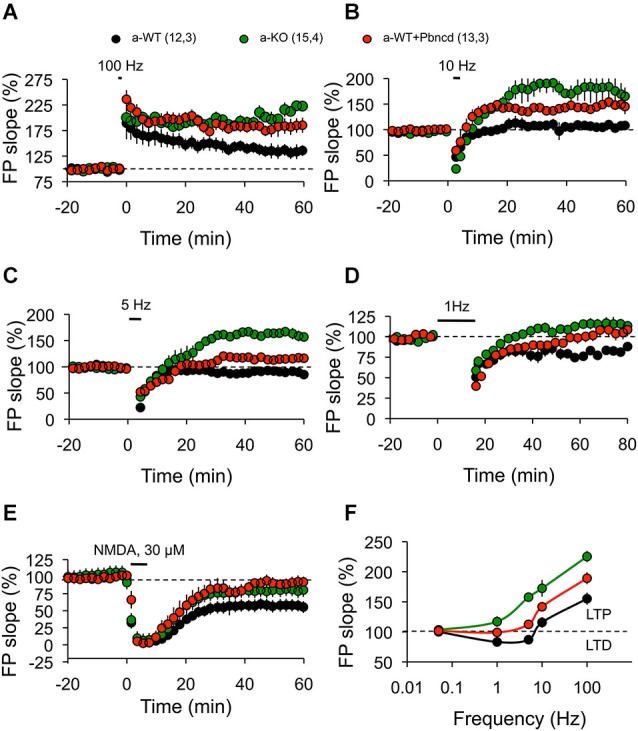
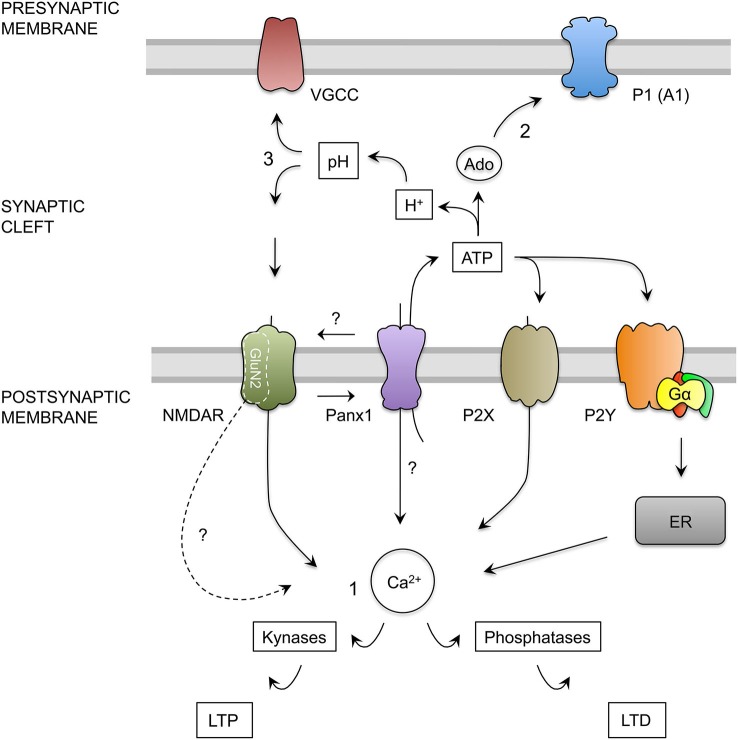
The threshold for bidirectional modification of synaptic plasticity is known to be controlled by several factors, including the balance between protein phosphorylation and dephosphorylation, postsynaptic free Ca(2+) concentration and NMDA receptor (NMDAR) composition of GluN2 subunits. Pannexin 1 (Panx1), a member of the integral membrane protein family, has been shown to form non-selective channels and to regulate the induction of synaptic plasticity as well as hippocampal-dependent learning. Although Panx1 channels have been suggested to play a role in excitatory long-term potentiation (LTP), it remains unknown whether these channels also modulate long-term depression (LTD) or the balance between both types of synaptic plasticity. To study how Panx1 contributes to excitatory synaptic efficacy, we examined the age-dependent effects of eliminating or blocking Panx1 channels on excitatory synaptic plasticity within the CA1 region of the mouse hippocampus. By using different protocols to induce bidirectional synaptic plasticity, Panx1 channel blockade or lack of Panx1 were found to enhance LTP, whereas both conditions precluded the induction of LTD in adults, but not in young animals. These findings suggest that Panx1 channels restrain the sliding threshold for the induction of synaptic plasticity and underlying brain mechanisms of learning and memory.
Keywords: LTD; LTP; NMDA receptors; hippocampus; mice; pannexin 1; synaptic plasticity.
Figures
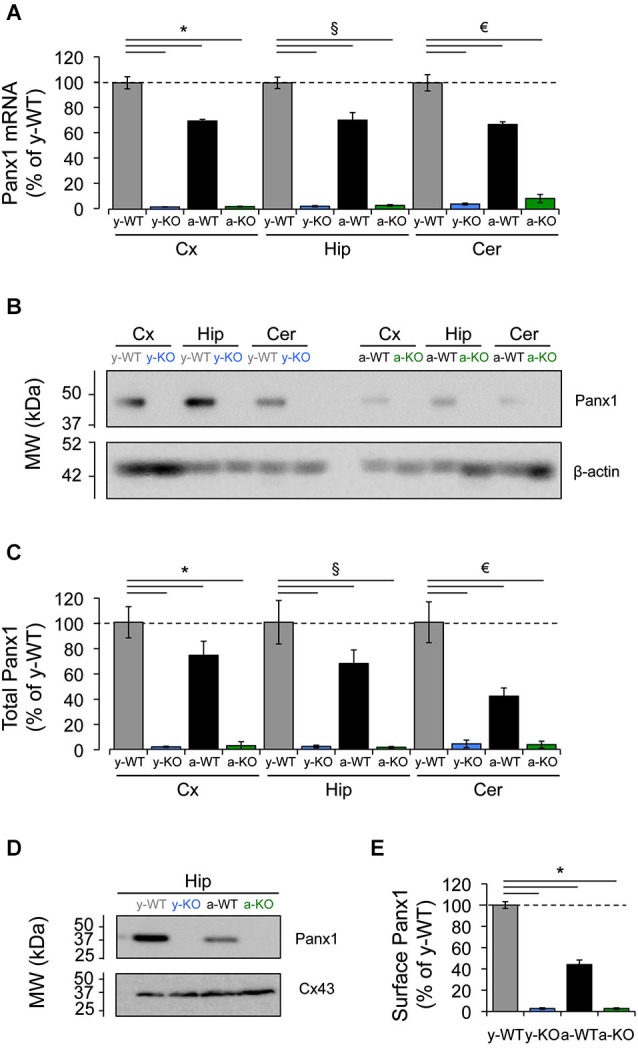
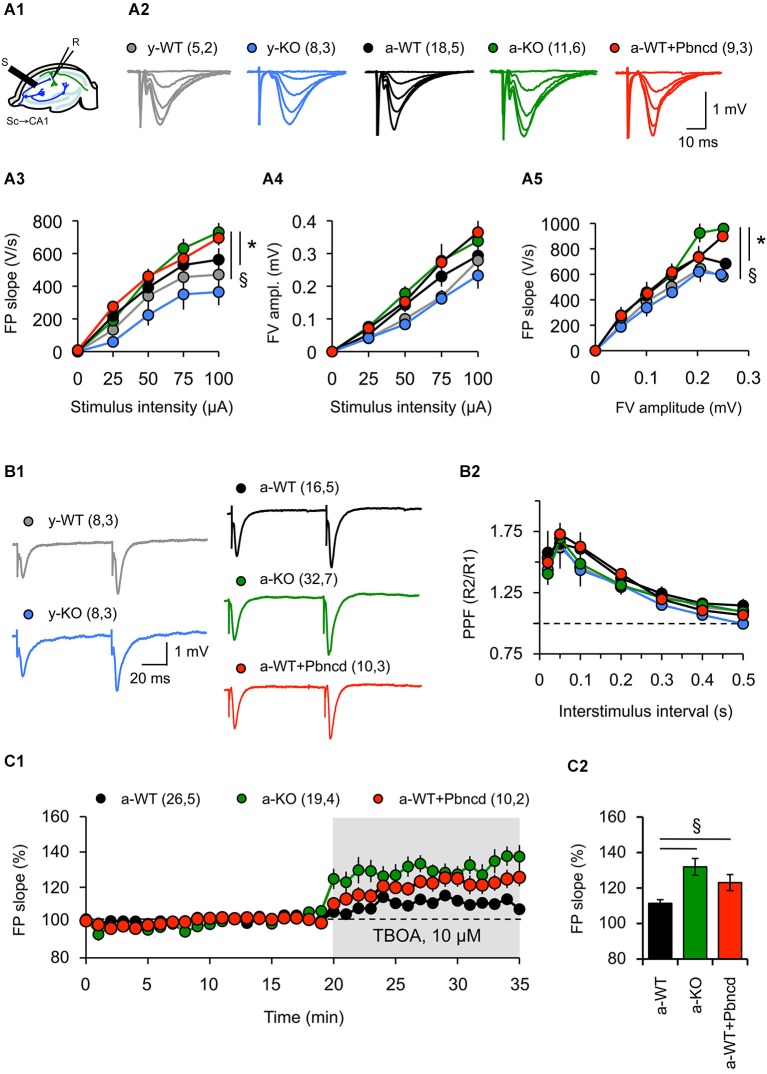
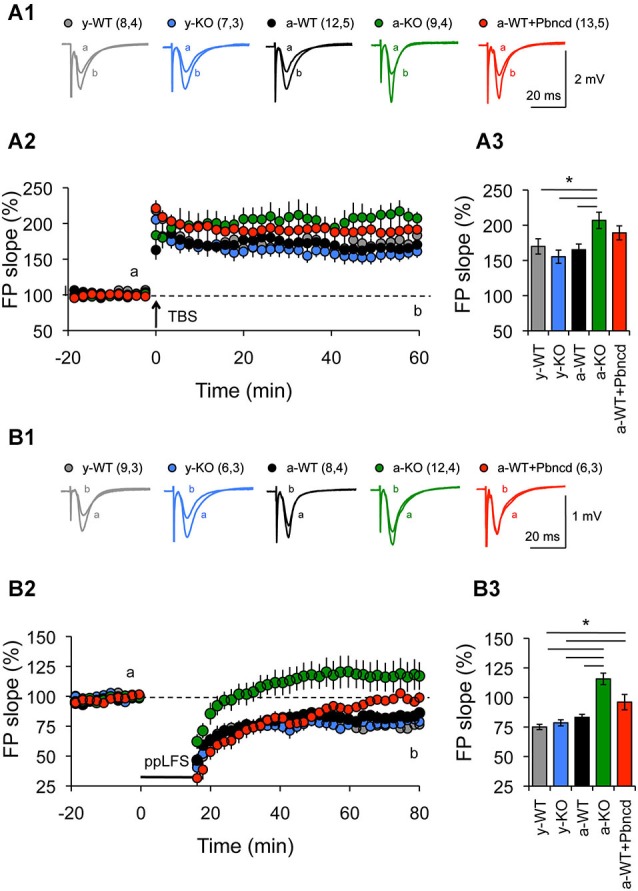
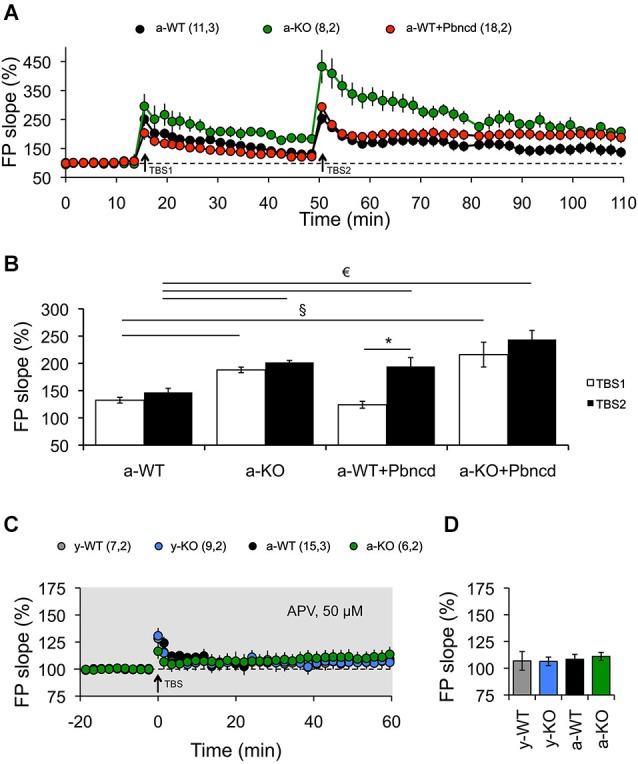
References
-
- Anselmi F., Hernandez V. H., Crispino G., Seydel A., Ortolano S., Roper S. D., et al. (2008). ATP release through connexin hemichannels and gap junction transfer of second messengers propagate Ca2+ signals across the inner ear. Proc. Natl. Acad. Sci. U S A 105, 18770–18775 10.1073/pnas.0800793105 - DOI - PMC - PubMed
-
- Ardiles A. O., Tapia-Rojas C. C., Mandal M., Alexandre F., Kirkwood A., Inestrosa N. C., et al. (2012). Post-synaptic dysfunction is associated with spatial and object recognition memory loss in a natural model of Alzheimer’s disease. Proc. Natl. Acad. Sci. U S A 109, 13835–13840 10.1073/pnas.1201209109 - DOI - PMC - PubMed
-
- Bartlett T. E., Bannister N. J., Collett V. J., Dargan S. L., Massey P. V., Bortolotto Z. A., et al. (2007). Differential roles of NR2A and NR2B-containing NMDA receptors in LTP and LTD in the CA1 region of two-week old rat hippocampus. Neuropharmacology 52, 60–70 10.1016/j.neuropharm.2006.07.013 - DOI - PubMed
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous