

Unfolded protein response-induced ERdj3 secretion links ER stress to extracellular proteostasis
- PMID: 25361606
- PMCID: PMC4291477
- DOI: 10.15252/embj.201488896
Unfolded protein response-induced ERdj3 secretion links ER stress to extracellular proteostasis
Abstract
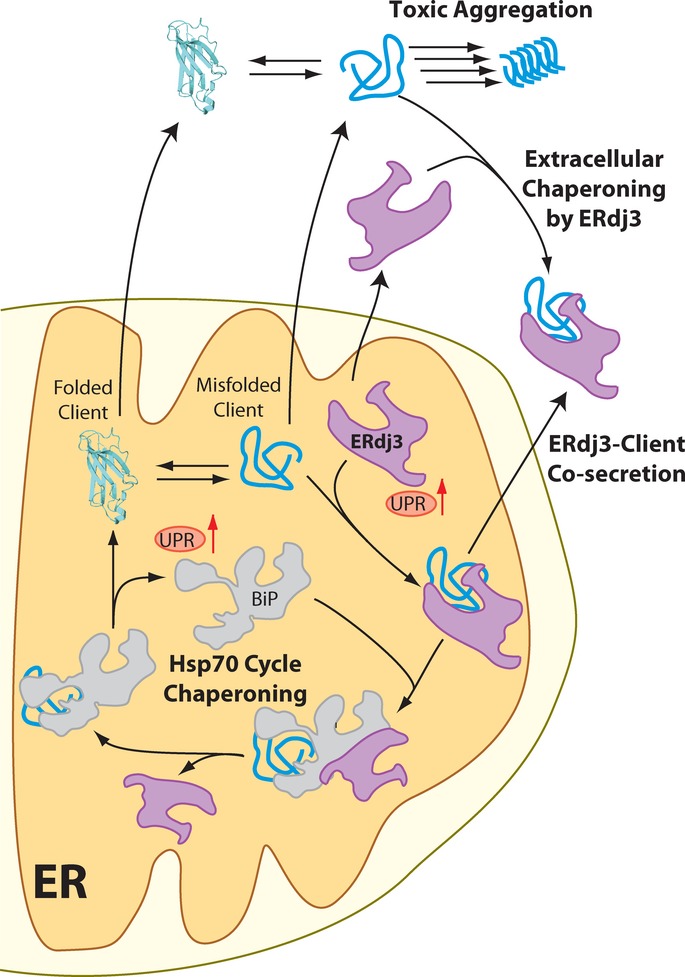
The Unfolded Protein Response (UPR) indirectly regulates extracellular proteostasis through transcriptional remodeling of endoplasmic reticulum (ER) proteostasis pathways. This remodeling attenuates secretion of misfolded, aggregation-prone proteins during ER stress. Through these activities, the UPR has a critical role in preventing the extracellular protein aggregation associated with numerous human diseases. Here, we demonstrate that UPR activation also directly influences extracellular proteostasis through the upregulation and secretion of the ER HSP40 ERdj3/DNAJB11. Secreted ERdj3 binds misfolded proteins in the extracellular space, substoichiometrically inhibits protein aggregation, and attenuates proteotoxicity of disease-associated toxic prion protein. Moreover, ERdj3 can co-secrete with destabilized, aggregation-prone proteins in a stable complex under conditions where ER chaperoning capacity is overwhelmed, preemptively providing extracellular chaperoning of proteotoxic misfolded proteins that evade ER quality control. This regulated co-secretion of ERdj3 with misfolded clients directly links ER and extracellular proteostasis during conditions of ER stress. ERdj3 is, to our knowledge, the first metazoan chaperone whose secretion into the extracellular space is regulated by the UPR, revealing a new mechanism by which UPR activation regulates extracellular proteostasis.
Keywords: ER stress; ERdj3; extracellular proteostasis; molecular chaperones; unfolded protein response.
© 2014 The Authors.
Figures
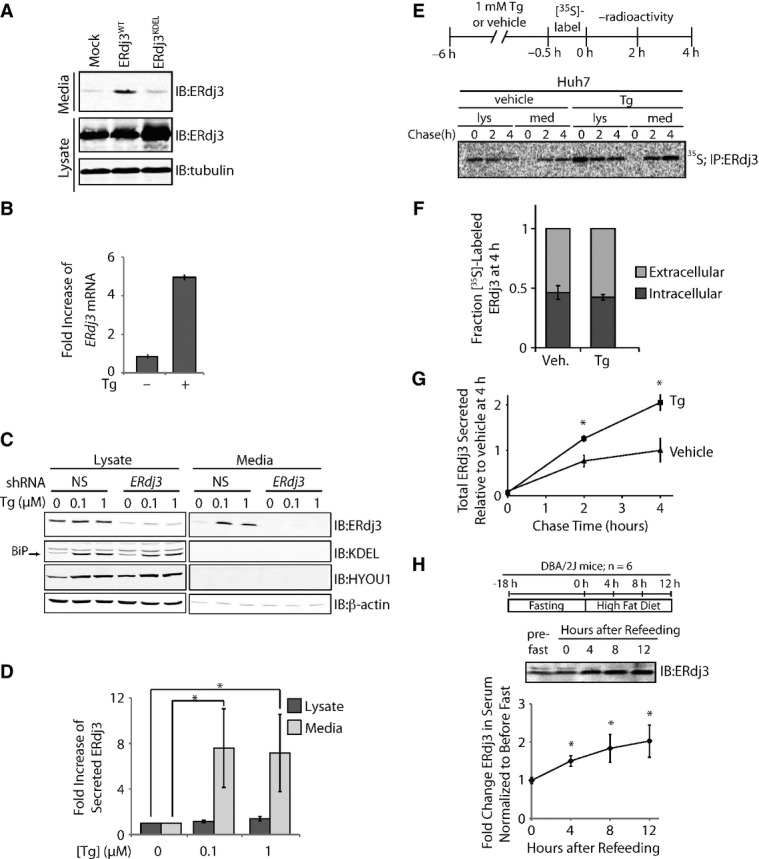
A Immunoblot of media and lysate collected from HEK293T cells overexpressing ERdj3WT or ERdj3KDEL. Fresh media was conditioned on cells for 24 h prior to harvest.
B qPCR analysis of ERdj3 mRNA in HEK293T cells treated with or without thapsigargin (Tg; 6 h, 100 nM). Error bars represent the mean ± 95% confidence interval as calculated in DataAssist 2.0 (n = 3).
C Representative immunoblot of lysates and media collected from HEK293T-Rex cells stably expressing non-silencing (NS) or ERdj3 shRNA. Cells were treated with the indicated concentration of Tg in fresh media for 16 h.
D Quantification of the fold increase of ERdj3 in lysates and media collected from HEK293T-Rex cells treated with the indicated concentration of Tg in fresh media for 16 h as in (C). *P < 0.05; n = 3, mean ± SEM.
E Representative autoradiogram of [35S]-labeled ERdj3 in lysates and media collected from Huh7 cells incubated in the presence or the absence of Tg (1 μM). The experimental paradigm for labeling is shown above.
F Quantification of fraction intracellular (dark gray) and extracellular (light gray) [35S]-labeled ERdj3 following a 4 h chase relative to total [35S]-labeled ERdj3 at 4 h (as shown in E). Error bars represent the standard error from biological replicates (n = 3).
G Quantification of total [35S]-labeled ERdj3 in media collected from cells pretreated with vehicle or Tg, as shown in (E). The media [35S]-labeled ERdj3 was measured by densitometry and is normalized to the amount of media [35S]-labeled ERdj3 at 4 h from vehicle-treated cells. *P-value < 0.05; n = 3, mean ± SEM.
H Representative immunoblot and quantification of sera collected at the indicated times from DBA/2J mice fed a high-fat (60% fat) diet after a 18 h fast. A schematic of the fasting/refeeding experimental setup is shown above. The quantification was normalized for each mouse to the amount of ERdj3 in the serum draw immediately prior the fast. *P-value < 0.05; n = 6, mean ± SEM.
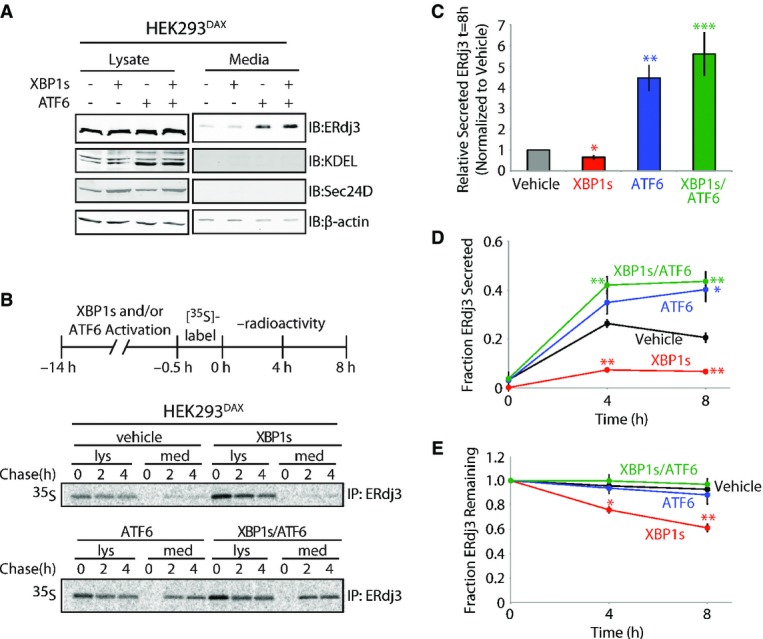
A Immunoblot of lysates and media collected from HEK293DAX cells following 16 h of activation of XBP1s (by 1 μg/ml dox) and/or ATF6 (by 10 μM TMP), as indicated (Shoulders et al, 2013). Immunoblots of the ATF6 target protein BiP (reactive with the KDEL antibody) and the XBP1s target Sec24D confirm the small-molecule activation of these transcription factors.
B Representative autoradiogram of [35S]-labeled ERdj3 immunopurified from lysates and media collected from HEK293DAX cells following preactivation of XBP1s (by 1 μg/ml dox) and/or ATF6 (by 10 μM TMP) (Shoulders et al, 2013). The experimental protocol is shown above.
C Quantification of relative amounts of [35S]-labeled ERdj3 in media at 8 h collected from cells treated as shown in (B) (n = 3). The media [35S]-labeled ERdj3 was measured by densitometry and is normalized to the amount of media [35S]-labeled ERdj3 at 8 h from vehicle-treated cells.
D, E Quantification of the fraction ERdj3 secreted (D) and the fraction ERdj3 remaining (E) from autoradiograms as shown in (B). The fractions of ERdj3 secreted/ERdj3 remaining were calculated by normalizing the recovered [35S]-labeled ERdj3 signal in the media and in the lysates at t = 4 or 8 h to the total amount of [35S]-labeled ERdj3 signal in the media and lysates at t = 0 h.
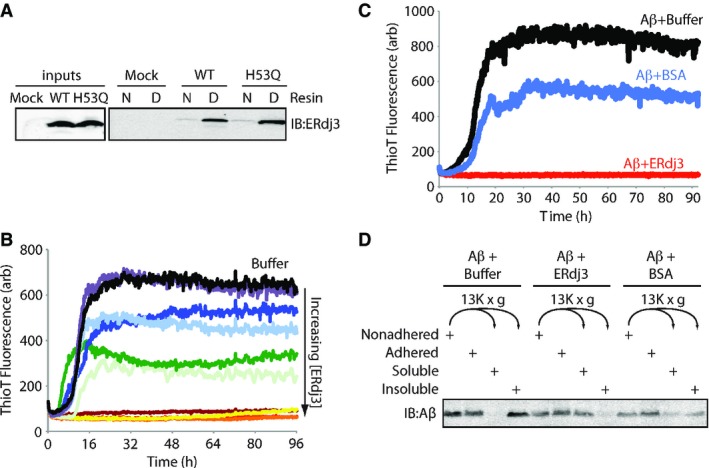
A Immunoblot of conditioned media collected from HEK293T cells transiently transfected with ERdj3WT or ERdj3H53Q and incubated with an affinity resin consisting of native (N) or guanidinium hydrochloride (GdnHCl)-denatured (D) RNase A covalently coupled to sepharose resin. Resin was incubated with media for 2 h at ambient temperature, followed by washing and then elution in Laemmli reducing buffer.
B Plot showing the time-dependent increase in ThioT fluorescence (ex: 440 nm, em: 485 nm) of Aβ1–40 incubated (10 μM, 37°C, pH 7.2) with regular agitation in the presence or the absence (black) of recombinant ERdj3 (RERdj3). The RERdj3 concentrations and molar ratios of RERdj3 to Aβ1–40 are as follows: violet: 1.5 nM, 1:6,561; light blue: 4.6 nM, 1:2,187; dark blue: 14 nM, 1:729; dark green: 41 nM, 1:243; light green: 120 nM, 1:81; yellow: 370 nM, 1:27; orange: 1.1 μM, 1:9; red: 3.3 μM, 1:3, or maroon: 10 μM, 1:1. All traces represent the average of three replicates.
C Plot showing the time-dependent increase in ThioT fluorescence of Aβ1–40 incubated (10 μM, 37°C, pH 7.2) with regular agitation in the presence or the absence of BSA (14 μg/ml) or RERdj3 (14 μg/ml; 370 nM, RERdj3: Aβ = 1:27). All traces represent the average of three replicates.
D Immunoblot of Aβ1–40 incubated (10 μM, 37°C, pH 7.2) with regular agitation for 90 h in the presence or the absence of RERdj3 (14 μg/ml; 370 nM, RERdj3: Aβ1–40 = 1:27) or BSA (14 μg/ml). The reaction solution in the well was collected (Nonadhered) and separated into soluble and insoluble fractions by centrifugation. These samples, as well as the washed wells of the plate (Adhered), were denatured in 8 M GdnHCl with sonication and analyzed by SDS-PAGE/immunoblotting.
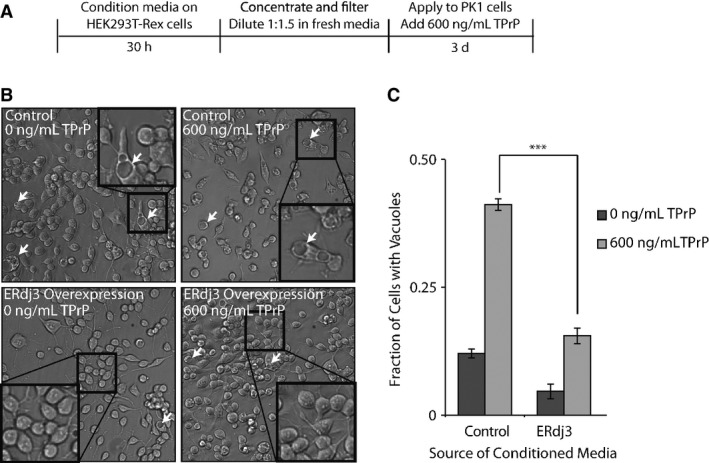
A Schematic illustrating the treatment of PK1 cells with ERdj3-conditioned media. HEK293T-Rex (Control) and HEK293T-Rex cells stably overexpressing tet-inducible ERdj3WT on poly-D-lysine-coated plates were treated with dox (1 μg/ml) overnight to induce ERdj3WT expression. The media was replaced with fresh OptiMEM I + 5% FBS, which was conditioned for 30 h, sterile-filtered, concentrated 5× against a 3 kD MWCO membrane, and diluted 1:1.5 with fresh OptiMEM I + 5% FBS. PK1 cells were plated in OptiMEM I + 5% FBS and after 24 h were treated with conditioned media and TPrP (600 ng/ml) for 3 days.
B Representative images under 136× magnification of PK1 cells incubated with control conditioned media collected from HEK293T-Rex or conditioned media prepared from HEK293T-Rex cells overexpressing ERdj3WT in the presence or the absence of TPrP (600 ng/ml), as described in (A). Arrows indicate representative cells containing vacuoles. Insets show 544× magnification of a typical region of the image.
C Quantification of vacuole formation from images as shown in (B). Equal area images comprising ∼1,200 cells per well were counted for each sample. Counting of cells and of cells with identifiable vacuoles was done using the Fiji image processing package (Schindelin et al, 2012). ***P-value < 0.001 (n = 3); mean ± SEM.
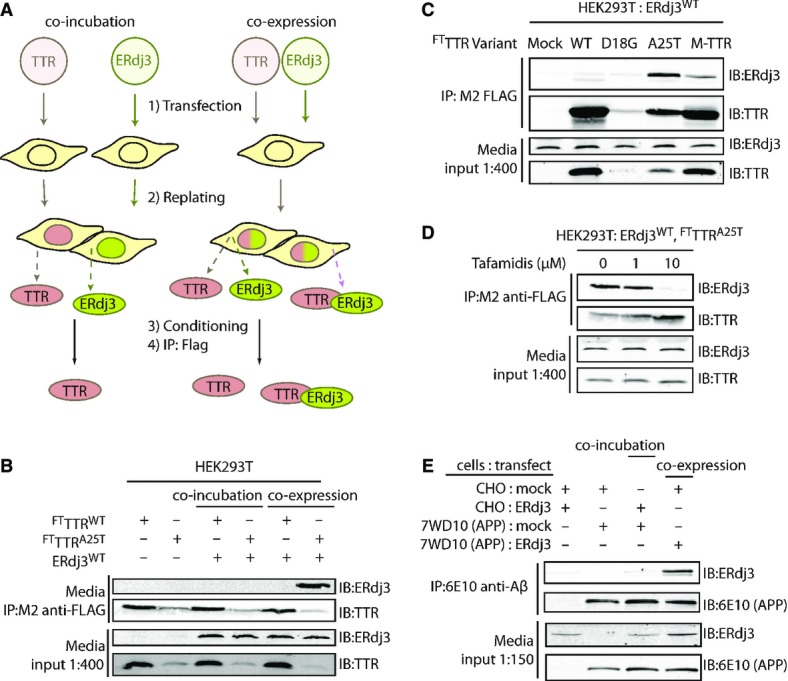
A Schematic describing the co-incubation and co-expression experiments used to demonstrate ERdj3 co-secretion with destabilized TTR. For co-incubation, HEK293T cells overexpressing FLAG-tag TTR (FTTTR) variants were seeded with an equal number of cells overexpressing ERdj3WT. For co-expression experiments, HEK293T cells are transfected with both FTTTR and ERdj3WT. FTTTR was immunoisolated with M2 anti-FLAG beads from media conditioned overnight on the cells, eluted, and separated by SDS-PAGE for immunoblotting.
B Immunoblot of M2 anti-FLAG immunopurifications from the conditioned media collected from HEK293T cells overexpressing FTTTRWT, FTTTRA25T, and/or ERdj3WT either as a co-incubation or co-expression experiment as shown in (A). Beads were washed in RIPA buffer prior to elution. Media inputs (1:400) are shown as a control.
C Immunoblot of M2 anti-FLAG immunopurifications from media collected from HEK293T cells overexpressing ERdj3WT and FTTTR variants as indicated. Media inputs (1:400) are shown as a control.
D Immunoblot of M2 anti-FLAG immunopurifications from conditioned media of HEK293T cells overexpressing FTTTRA25T and ERdj3WT in the presence of the indicated concentrations of Tafamidis, a kinetic stabilizer of TTR tetramers. Media inputs (1:400) are shown as a control.
E Immunoblot of 6E10 anti-APP immunopurifications from the conditioned media of CHO cells or CHO-derived APP751-expressing (7WD10) cells overexpressing ERdj3WT as indicated. Cells were replated together as indicated 24 h after transfection at equal stoichiometry, and media conditioned for 36 h. In this case, ERdj3WT-transfected CHO cells replated with mock-transfected 7WD10 cells serves as the co-incubation condition, while mock-transfected CHO cells replated with ERdj3WT-transfected 7WD10 cells comprise the co-expression condition. Media inputs (1:150) are shown as a control.
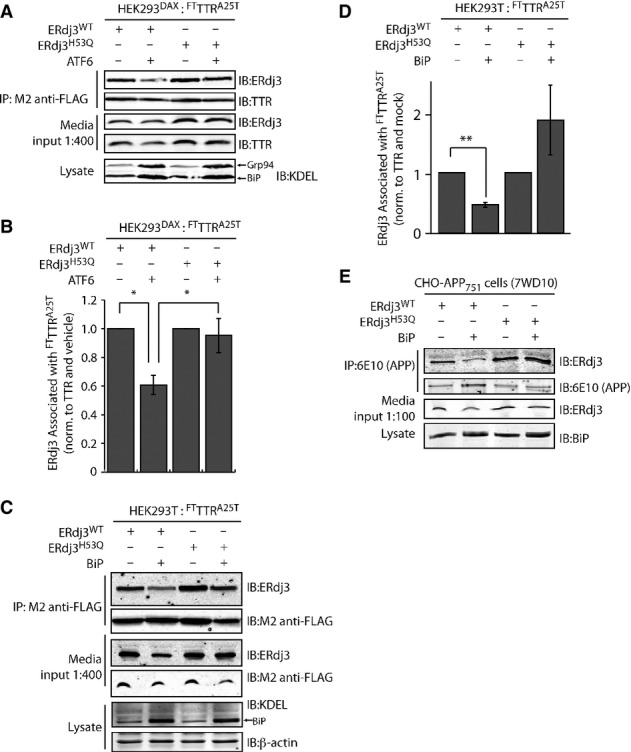
A Representative immunoblot of M2 anti-FLAG immunopurification of media conditioned for 24 h on HEK293DAX cells co-overexpressing FTTTRA25T and ERdj3WT or ERdj3H53Q. Media contained vehicle or trimethoprim (TMP, 10 μM, 16 h; activates ATF6), as indicated. Media inputs (1:400) are shown as a control. Lysates from these cells are also shown to confirm the increase of the ATF6 target proteins BiP and GRP94 in cells following TMP-dependent activation of ATF6.
B Bar graph depicting quantification of the change in the amount of ERdj3 co-immunoprecipitating with TTRA25T following activation of ATF6 as in (A). The ERdj3/TTR ratio is normalized to the ratio in the absence of ATF6 activation. *P-value < 0.05; n = 3, mean ± SEM.
C Representative immunoblot of M2 anti-FLAG immunopurification of media conditioned for 24 h on HEK293T cells co-overexpressing FTTTRA25T, ERdj3WT, ERdj3H53Q, and/or BiP as indicated. Media inputs (1:400) are shown as a control. Lysates from these cells are also shown to confirm the increase of BiP afforded by overexpression.
D Quantification of immunoblots of M2 anti-FLAG immunopurification of media collected from cells co-overexpressing either ERdj3WT or ERdj3H53Q, FTTTRA25T, and BiP as in (C). The ERdj3/TTR ratio is normalized to the ratio in the absence of BiP overexpression. **P-value < 0.01; n = 5, mean ± SEM.
E Immunoblot of 6E10 anti-APP immunopurifications from the conditioned media of CHO-derived APP751-expressing (7WD10) cells overexpressing ERdj3WT, ERdj3H53Q, and BiP as indicated. Media inputs (1:100) are shown as a control. Lysates from these cells are also shown to confirm the increase of BiP afforded by overexpression.
References
-
- Adachi Y, Yamamoto K, Okada T, Yoshida H, Harada A, Mori K. ATF6 is a transcription factor specializing in the regulation of quality control proteins in the endoplasmic reticulum. Cell Struct Funct. 2008;33:75–89. - PubMed
-
- Aguzzi A, Heikenwalder M, Polymenidou M. Insights into prion strains and neurotoxicity. Nat Rev Mol Cell Biol. 2007;8:552–561. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- R01DK102635/DK/NIDDK NIH HHS/United States
- R21 NS079882/NS/NINDS NIH HHS/United States
- R21NS079882/NS/NINDS NIH HHS/United States
- R01HL052173/HL/NHLBI NIH HHS/United States
- R37 DK042394/DK/NIDDK NIH HHS/United States
- R37DK042394/DK/NIDDK NIH HHS/United States
- F32-HL099245/HL/NHLBI NIH HHS/United States
- R21NS081519/NS/NINDS NIH HHS/United States
- R01 HL052173/HL/NHLBI NIH HHS/United States
- R01 DK102635/DK/NIDDK NIH HHS/United States
- R01 DK088227/DK/NIDDK NIH HHS/United States
- F32 HL099245/HL/NHLBI NIH HHS/United States
- R37 DK046335/DK/NIDDK NIH HHS/United States
- R37DK046335/DK/NIDDK NIH HHS/United States
- R21 NS081519/NS/NINDS NIH HHS/United States
- R01DK088227/DK/NIDDK NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
