

Transcriptome analysis reveals novel regulatory mechanisms in a genome-reduced bacterium
- PMID: 25361977
- PMCID: PMC4245973
- DOI: 10.1093/nar/gku976
Transcriptome analysis reveals novel regulatory mechanisms in a genome-reduced bacterium
Abstract
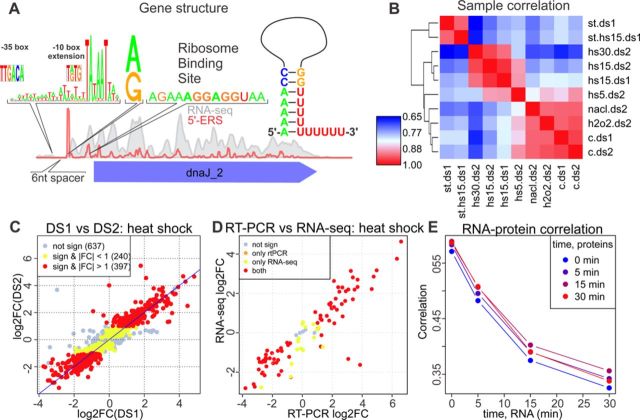
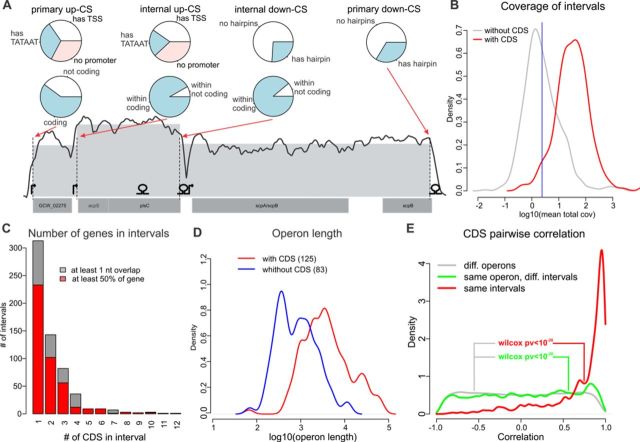
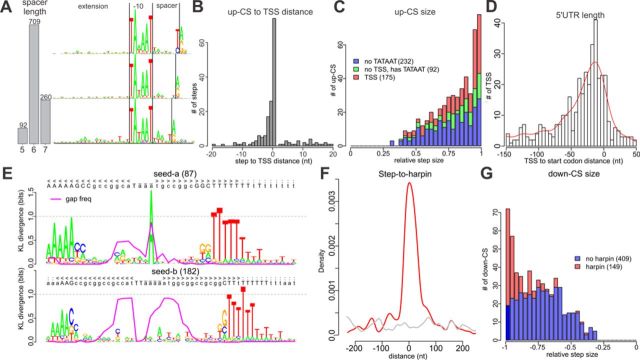
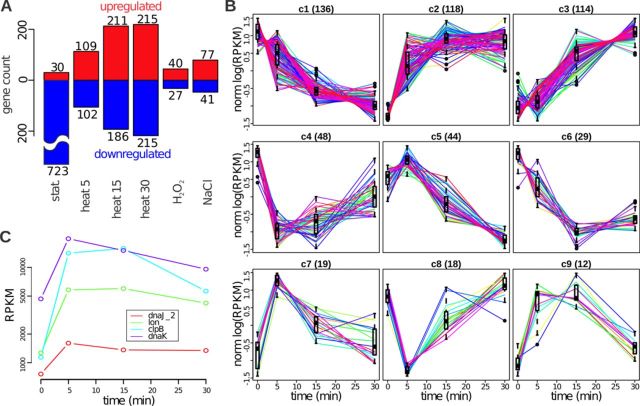
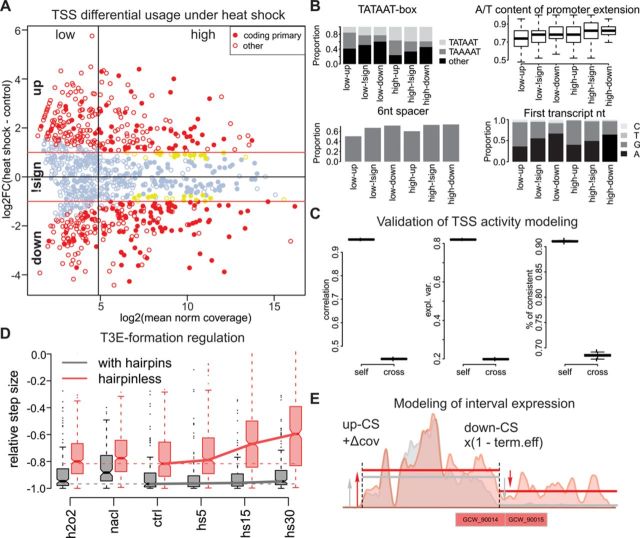
The avian bacterial pathogen Mycoplasma gallisepticum is a good model for systems studies due to small genome and simplicity of regulatory pathways. In this study, we used RNA-Seq and MS-based proteomics to accurately map coding sequences, transcription start sites (TSSs) and transcript 3'-ends (T3Es). We used obtained data to investigate roles of TSSs and T3Es in stress-induced transcriptional responses. We identified 1061 TSSs at a false discovery rate of 10% and showed that almost all transcription in M. gallisepticum is initiated from classic TATAAT promoters surrounded by A/T-rich sequences. Our analysis revealed the pronounced operon structure complexity: on average, each coding operon has one internal TSS and T3Es in addition to the primary ones. Our transcriptomic approach based on the intervals between the two nearest transcript ends allowed us to identify two classes of T3Es: strong, unregulated, hairpin-containing T3Es and weak, heat shock-regulated, hairpinless T3Es. Comparing gene expression levels under different conditions revealed widespread and divergent transcription regulation in M. gallisepticum. Modeling suggested that the core promoter structure plays an important role in gene expression regulation. We have shown that the heat stress activation of cryptic promoters combined with the hairpinless T3Es suppression leads to widespread, seemingly non-functional transcription.
© The Author(s) 2014. Published by Oxford University Press on behalf of Nucleic Acids Research.
Figures
References
-
- Levisohn S., Kleven S.H. Avian mycoplasmosis (Mycoplasma gallisepticum) Rev. sci. tech. Off. int. Epiz. 2000;19:425–442. - PubMed
-
- Chang L.-J., Chen W.-H., Minion F.C., Shiuan D. Mycoplasmas regulate the expression of heat-shock protein genes through CIRCE-HrcA interactions. Biochemical and biophysical research communications. 2008;367:213–218. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
