

Intracellular metabolite levels shape sulfur isotope fractionation during microbial sulfate respiration
- PMID: 25362045
- PMCID: PMC4280625
- DOI: 10.1073/pnas.1407502111
Intracellular metabolite levels shape sulfur isotope fractionation during microbial sulfate respiration
Abstract
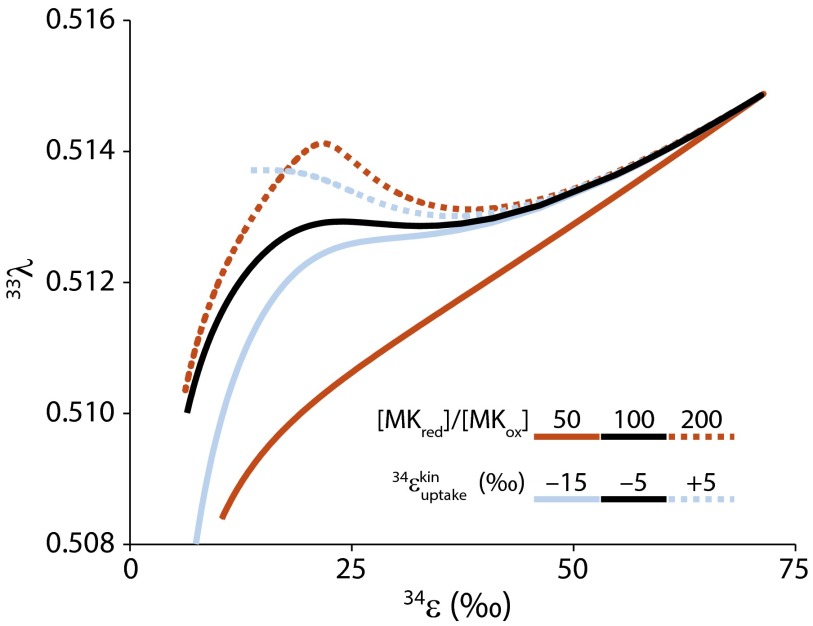
We present a quantitative model for sulfur isotope fractionation accompanying bacterial and archaeal dissimilatory sulfate respiration. By incorporating independently available biochemical data, the model can reproduce a large number of recent experimental fractionation measurements with only three free parameters: (i) the sulfur isotope selectivity of sulfate uptake into the cytoplasm, (ii) the ratio of reduced to oxidized electron carriers supporting the respiration pathway, and (iii) the ratio of in vitro to in vivo levels of respiratory enzyme activity. Fractionation is influenced by all steps in the dissimilatory pathway, which means that environmental sulfate and sulfide levels control sulfur isotope fractionation through the proximate influence of intracellular metabolites. Although sulfur isotope fractionation is a phenotypic trait that appears to be strain specific, we show that it converges on near-thermodynamic behavior, even at micromolar sulfate levels, as long as intracellular sulfate reduction rates are low enough (<<1 fmol H2S⋅cell(-1)⋅d(-1)).
Keywords: dissimilatory sulfate reduction; flux–force relationship; sulfur isotope fractionation.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
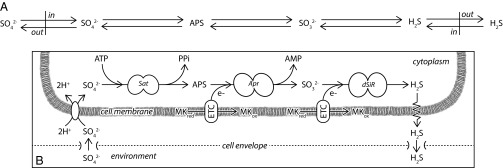
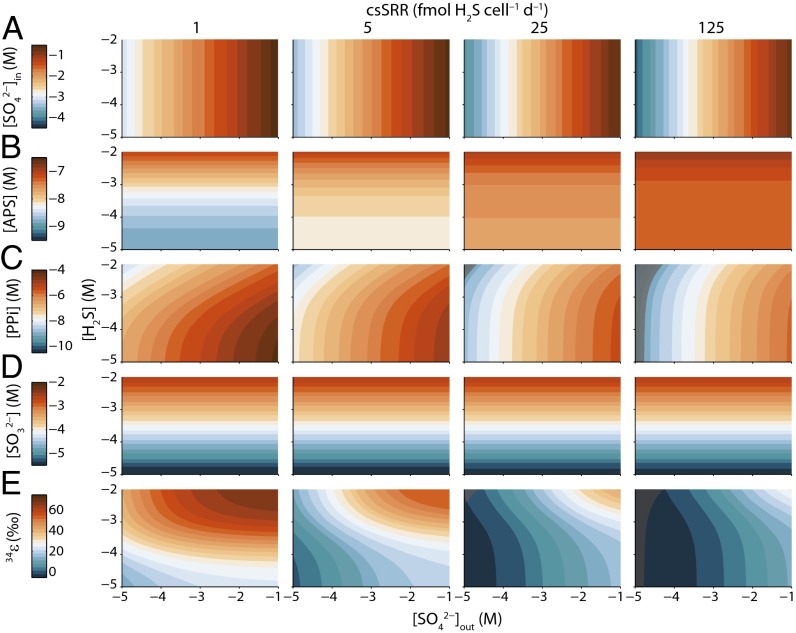
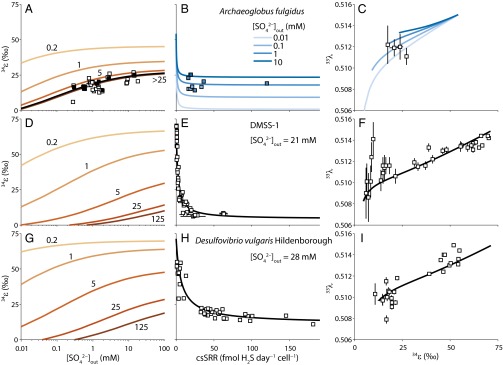
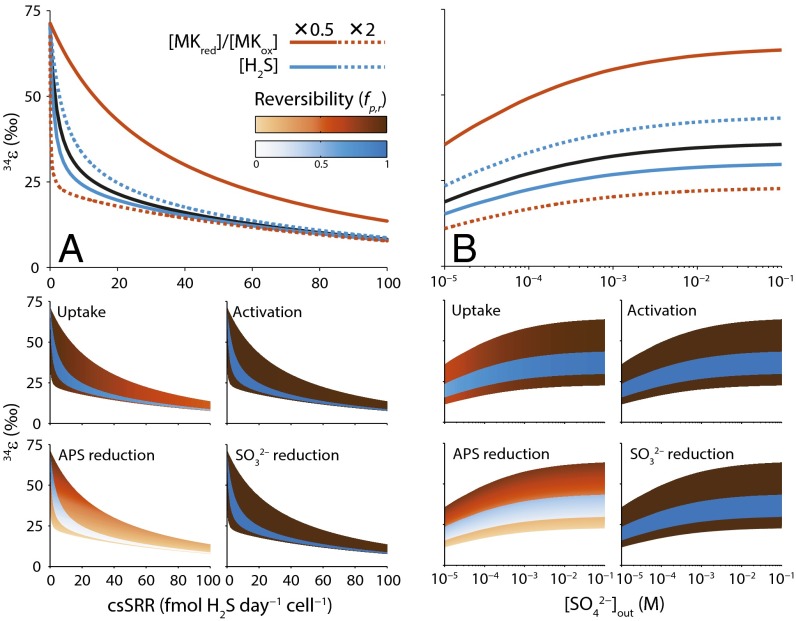
Comment in
-
Predictive isotope model connects microbes in culture and nature.Proc Natl Acad Sci U S A. 2014 Dec 23;111(51):18102-3. doi: 10.1073/pnas.1420670111. Epub 2014 Dec 5. Proc Natl Acad Sci U S A. 2014. PMID: 25480549 Free PMC article. No abstract available.
References
-
- Rabus R, Hansen T, Widdel F. In: Dissimilatory Sulfate- and Sulfur-Reducing Prokaryotes. Rosenberg E, DeLong E, Lory S, Stackebrandt E, Thompson F, editors. Springer; Berlin: 2013. pp. 309–404.
-
- Thode H, Kleerekoper H, McElcheran D. Isotope fractionation in the bacterial reduction of sulphate. Research. 1951;4:581–582.
-
- Harrison A, Thode H. Mechanism of the bacterial reduction of sulphate from isotope fractionation studies. Trans Faraday Soc. 1958;54:84–92.
-
- Kaplan IR, Rittenberg SC. Microbiological fractionation of sulphur isotopes. J Gen Microbiol. 1964;34(2):195–212. - PubMed
-
- Chambers LA, Trudinger PA, Smith JW, Burns MS. Fractionation of sulfur isotopes by continuous cultures of Desulfovibrio desulfuricans. Can J Microbiol. 1975;21(10):1602–1607. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
