

Dysregulation of autophagy in human follicular lymphoma is independent of overexpression of BCL-2
- PMID: 25362242
- PMCID: PMC4294376
- DOI: 10.18632/oncotarget.2605
Dysregulation of autophagy in human follicular lymphoma is independent of overexpression of BCL-2
Abstract
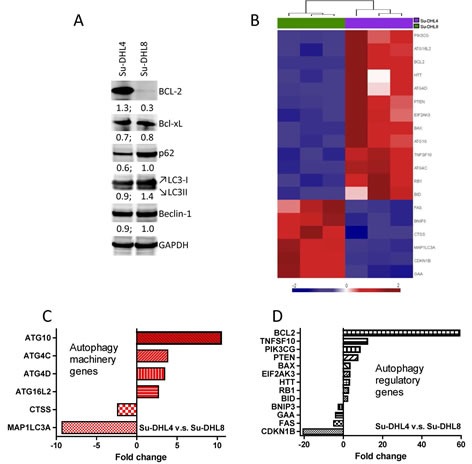
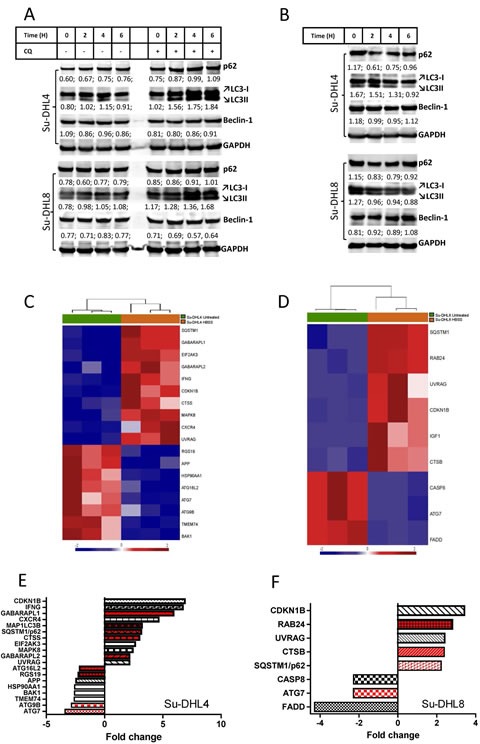
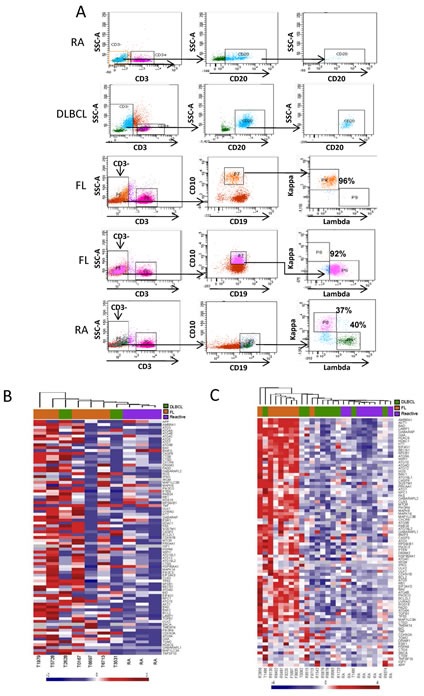
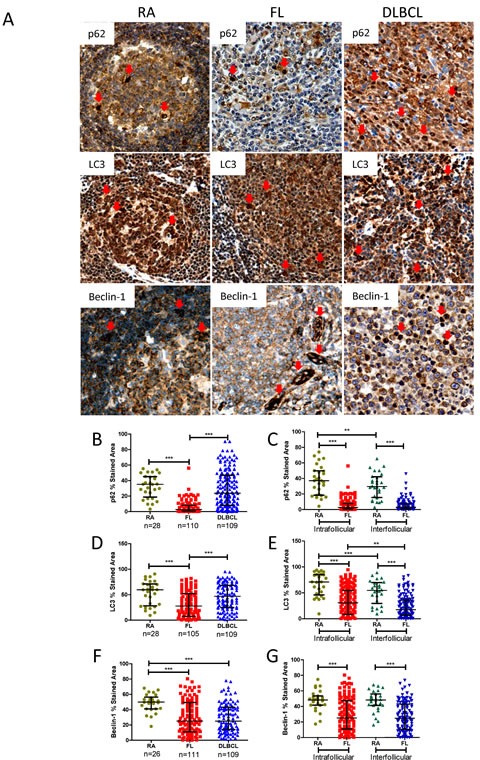
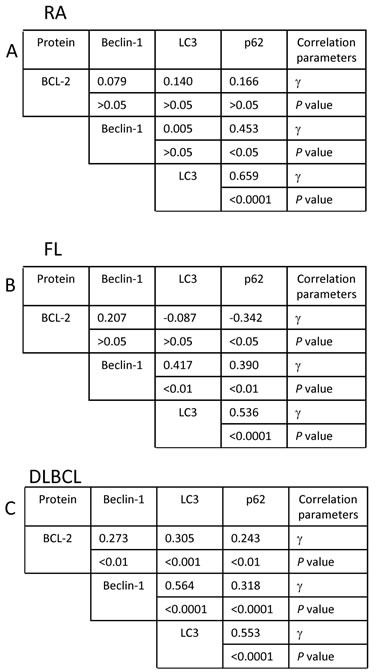
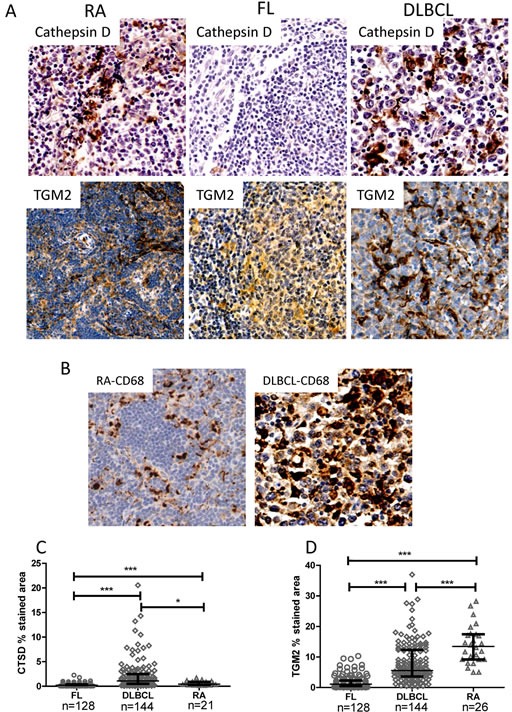
Overexpression of the anti-apoptotic protein BCL-2 is characteristic of human follicular lymphoma (FL) and some cases of diffuse large B cell lymphoma (DLBCL). We aimed to determine autophagy status in primary FL and DLBCL samples and the BCL-2+/BCL-2- lymphoma cell lines using both autophagy PCR array and tissue microarray (TMA). A greater number of autophagy machinery genes were up-regulated in the BCL-2+ Su-DHL4 cell line compared with BCL-2- Su-DHL8 cells, at both the basal level and in response to autophagic stress. The autophagy-related gene expression profiles were determined in purified and unpurified malignant human lymph node biopsies. Seven autophagy machinery genes were up-regulated in purified FL B-cells compared with reactive B-cells. Only 2 autophagy machinery genes were up-regulated in DLBCL B-cells. In unpurified tissue biopsies, 20 of 46 genes in FL and 2 of 5 genes in DLBCL with increased expression were autophagy machinery genes. Expression of autophagy substrates p62 and LC3 were determined by TMAs. FL samples showed significantly decreased levels of both p62 and LC3 compared with reactive and DLBCL, indicative of an increased autophagy activity in FL. In summary, these results demonstrate that FL showed increased basal autophagy activity, regardless of overexpression of BCL-2 in this disease.
Conflict of interest statement
The authors declare that they have no conflict of interest.
Figures
References
-
- Jia L, Dourmashkin RR, Allen PD, Gray AB, Newland AC, Kelsey SM. Inhibition of autophagy abrogates tumour necrosis factor alpha induced apoptosis in human T-lymphoblastic leukaemic cells. Br J Haematol. 1997;98(3):673–685. - PubMed
-
- Rao S, Tortola L, Perlot T, Wirnsberger G, Novatchkova M, Nitsch R, Sykacek P, Frank L, Schramek D, Komnenovic V, Sigl V, Aumayr K, Schmauss G, Fellner N, Handschuh S, Glosmann M, et al. A dual role for autophagy in a murine model of lung cancer. Nat Commun. 2014;5:3056. - PubMed
-
- Wu WK, Coffelt SB, Cho CH, Wang XJ, Lee CW, Chan FK, Yu J, Sung JJ. The autophagic paradox in cancer therapy. Oncogene. 2012;31(8):939–953. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
