

The genetic basis of chronic mountain sickness
- PMID: 25362634
- PMCID: PMC4280153
- DOI: 10.1152/physiol.00008.2014
The genetic basis of chronic mountain sickness
Abstract
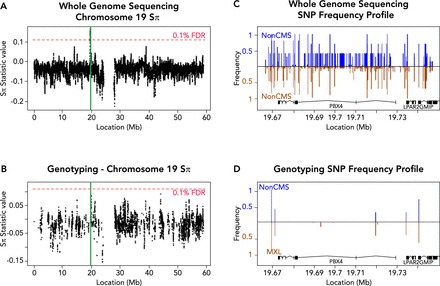
Chronic mountain sickness (CMS) is a disease that affects many high-altitude dwellers, particularly in the Andean Mountains in South America. The hallmark symptom of CMS is polycythemia, which causes increased risk of pulmonary hypertension and stroke (among other symptoms). A prevailing hypothesis in high-altitude medicine is that CMS results from a population-specific "maladaptation" to the hypoxic conditions at high altitude. In contrast, the prevalence of CMS is very low in other high-altitude populations (e.g., Tibetans and Ethiopians), which are seemingly well adapted to hypoxia. In recent years, concurrent with the advent of genomic technologies, several studies have investigated the genetic basis of adaptation to altitude. These studies have identified several candidate genes that may underlie the adaptation, or maladaptation. Interestingly, some of these genes are targeted by known drugs, raising the possibility of new treatments for CMS and other ischemic diseases. We review recent discoveries, alongside the methodologies used to obtain them, and outline some of the challenges remaining in the field.
©2014 Int. Union Physiol. Sci./Am. Physiol. Soc.
Conflict of interest statement
No conflicts of interest, financial or otherwise, are declared by the author(s).
Figures


References
-
- Beall CM. Andean, Tibetan, and Ethiopian patterns of adaptation to high-altitude hypoxia. Integr Comp Biol 46: 18–24, 2006. - PubMed
-
- Beall CM. Tibetan and Andean contrasts in adaptation to high-altitude hypoxia. Adv Exp Med Biol 475: 63–74, 2000. - PubMed
-
- Beall CM, Brittenham GM, Strohl KP, Blangero J, Williams-Blangero S, Goldstein MC, Decker MJ, Vargas E, Villena M, Soria R, Alarcon AM, Gonzales C. Hemoglobin concentration of high-altitude Tibetans and Bolivian Aymara. Am J Phys Anthropol 106: 385–400, 1998. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources