

Evolution of Metallo-β-lactamases: Trends Revealed by Natural Diversity and in vitro Evolution
- PMID: 25364574
- PMCID: PMC4212336
- DOI: 10.3390/antibiotics3030285
Evolution of Metallo-β-lactamases: Trends Revealed by Natural Diversity and in vitro Evolution
Abstract
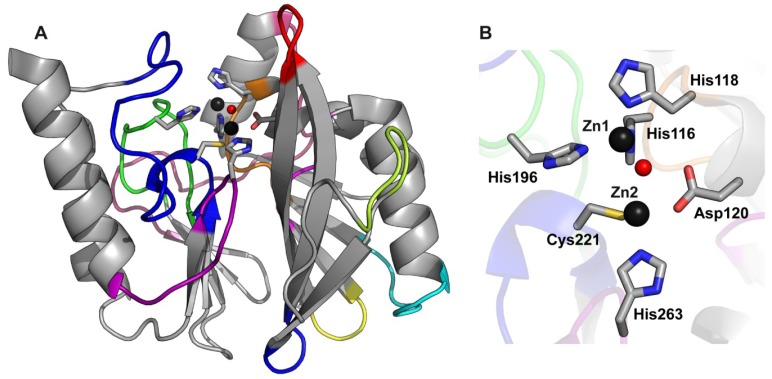
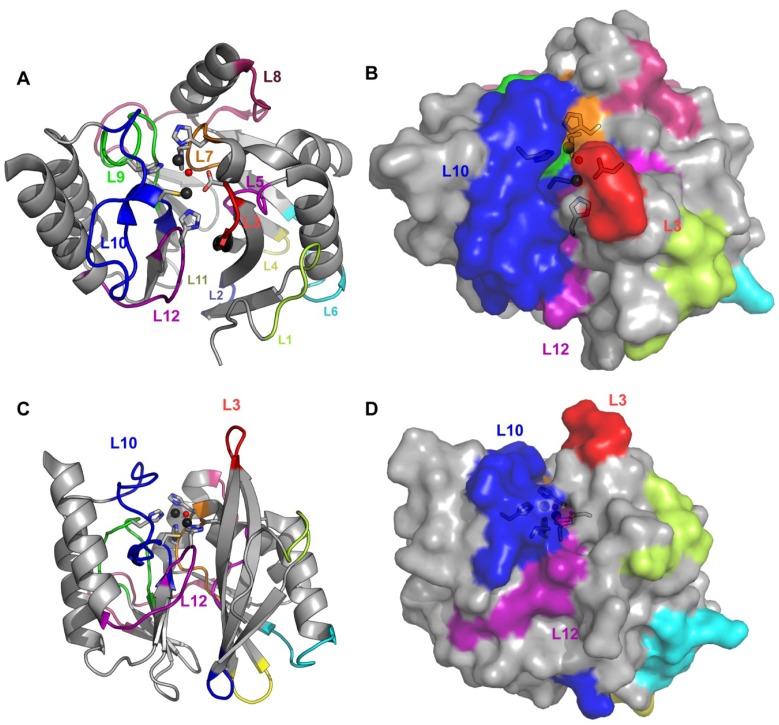
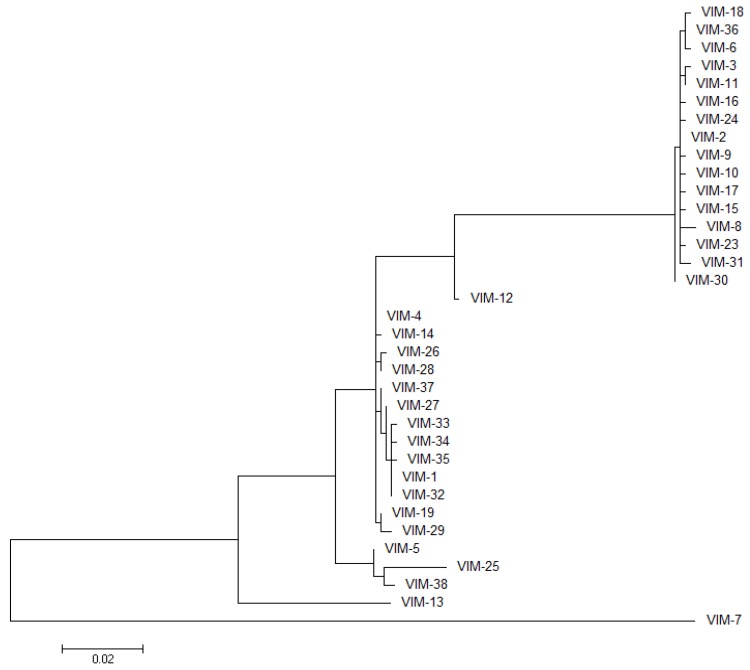
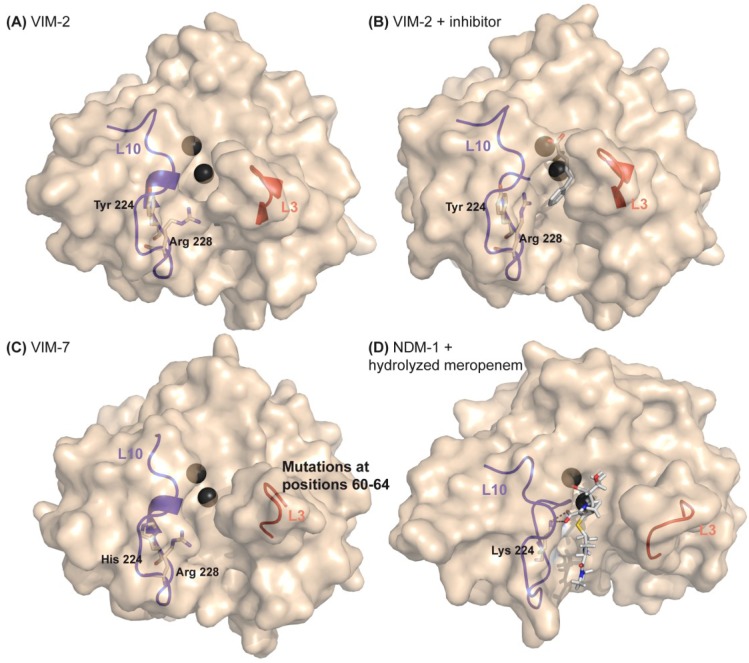
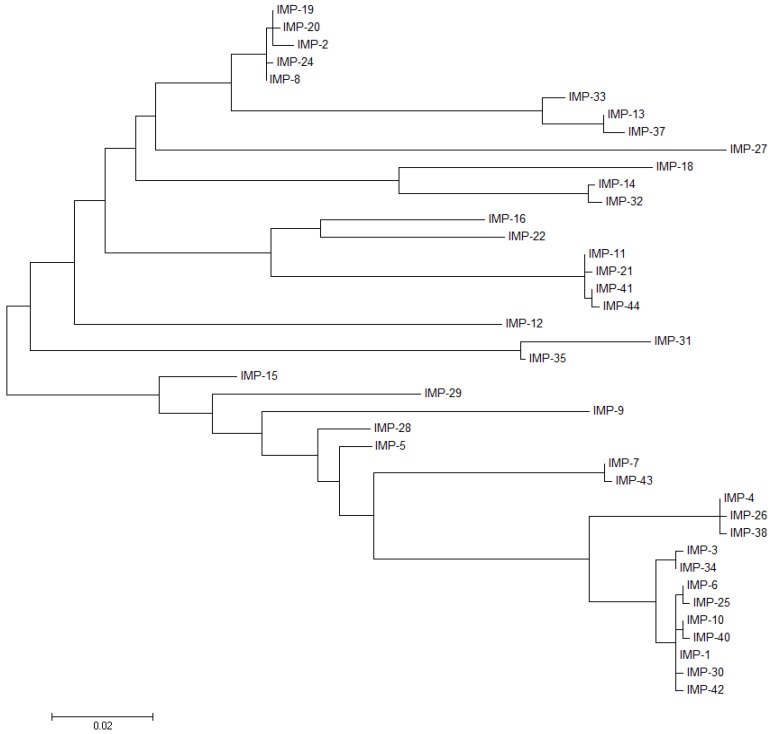
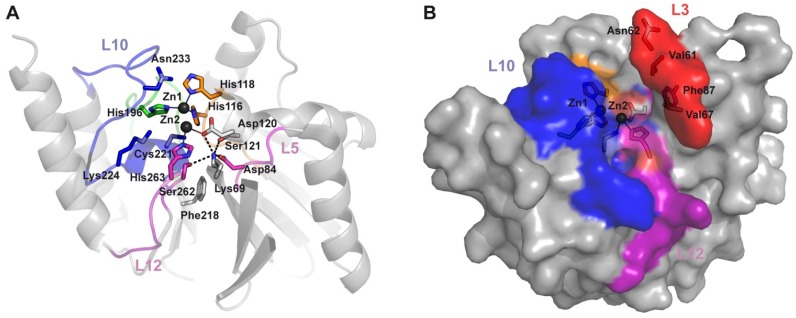
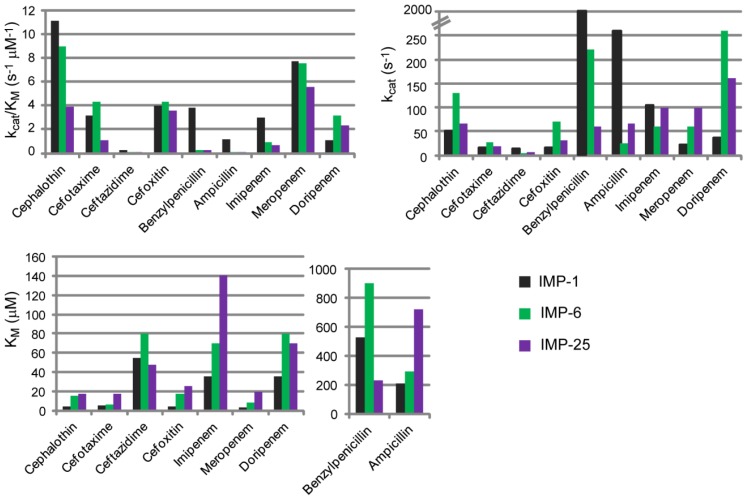
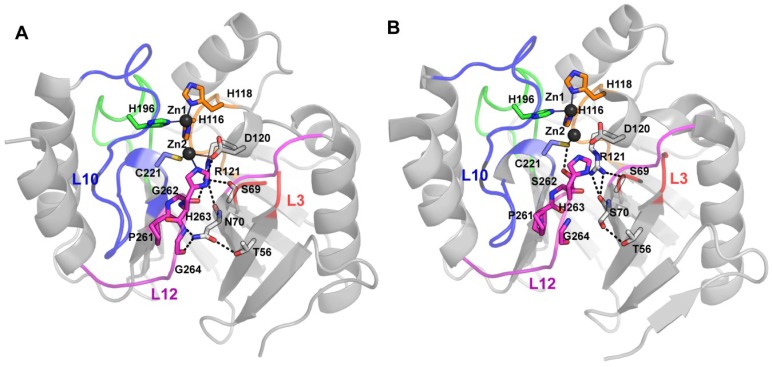
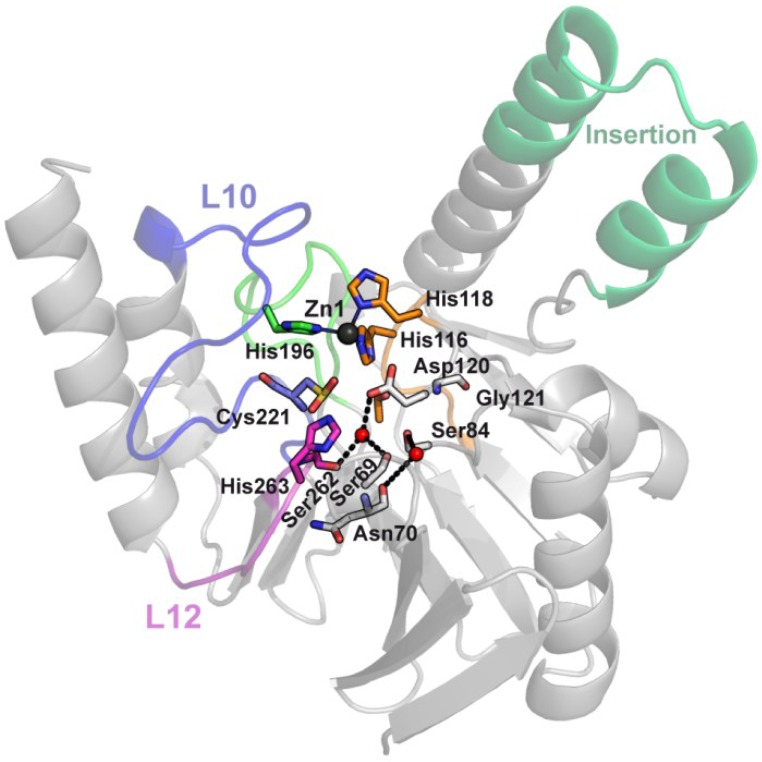
The production of β-lactamase enzymes is one of the most distributed resistance mechanisms towards β-lactam antibiotics. Metallo-β-lactamases constitute a worrisome group of these kinds of enzymes, since they present a broad spectrum profile, being able to hydrolyze not only penicillins, but also the latest generation of cephalosporins and carbapenems, which constitute at present the last resource antibiotics. The VIM, IMP, and NDM enzymes comprise the main groups of clinically relevant metallo-β-lactamases. Here we present an update of the features of the natural variants that have emerged and of the ones that have been engineered in the laboratory, in an effort to find sequence and structural determinants of substrate preferences. This knowledge is of upmost importance in novel drug design efforts. We also discuss the advances in knowledge achieved by means of in vitro directed evolution experiments, and the potential of this approach to predict natural evolution of metallo-β-lactamases.
Keywords: evolution; metallo-β-lactamases; resistance.
Figures



References
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous