

Review
doi: 10.1007/s12223-014-0359-6.
Epub 2014 Nov 1.
DNA repair mechanisms and human cytomegalovirus (HCMV) infection
Affiliations
- PMID: 25366712
- PMCID: PMC4429022
- DOI: 10.1007/s12223-014-0359-6
Item in Clipboard
Review
DNA repair mechanisms and human cytomegalovirus (HCMV) infection
Folia Microbiol (Praha).
2015 May.
Abstract
Herpesvirus infections, such as those induced by human cytomegalovirus (HCMV), induce specific DNA damages. DNA damages can lead to cell mutation, death, apoptosis and immune system activation. Various types of DNA damage are repaired through multiple repair pathways, such as base excision, nucleotide excision, homologous recombination and nonhomologous end joining. Changes in the activity of DNA repair proteins during viral infection can cause disturbances in the DNA repair system and change its mechanisms. This report reviews results from studies, assaying a DNA repair system in HCMV infection.
Figures

The base excision repair (BER) system. BER is initiated by a specific DNA glycosylases, which recognize and remove damaged or incorrect bases, creating AP sites. The AP endonucleases cleave an AP site to yield a 3′ hydroxyl adjacent to a 5′ deoxyribosephosphate (dRP). During short-patch BER, 5′dRP is displaced by DNA polymerase β (Polβ), which inserts a single nucleotide. Polβ is also involved in long-patch BER inserting the first nucleotide at reduced AP sites. The ligation step is performed by DNA ligases I and III. Ligase I interacts with Polβ and participates mainly in long-patch BER. DNA ligase III interacts with X-ray repair cross-complementing protein 1 (XRCC1) and Polβ and is involved only in short-patch BER

The nucleotide excision repair. a Global genomic repair (GGR), xeroderma pigmentosum, complementation group C (XPC), UV excision repair protein RAD23 homolog B (HR23B), replication protein A (RPA), xeroderma pigmentosum group A (XPA) or DNA damage-binding protein 1 and 2 (DDB1–DDB2) complex recognition DNA damage. DNA repair is performed by the transcription factor TFIIH, and excision of the lesion by xeroderma pigmentosum, complementation group G and F (XPG and XPF), excision repair cross-complementing 1 (ERCC1). Finally, resynthesis occurs by polymerase (Polδ or Polϵ) and ligation by DNA ligase I. b Transcription-coupled repair (TCR). First step, RNA polII (RNAPII) is blocked. This leads to assembly of Cockayne syndrome group A/B proteins (CSA, CSB) and/or the transcription elongation factor TFIIS at the site of the lesion, by which RNAPII is removed from the DNA or displaced from the lesion, making it accessible to the exonucleases XPF. Second step, XPG cleaving the lesion-containing DNA strand. Third step, resynthesis again occurs by Polδ or Polϵ and ligation by DNA ligase I
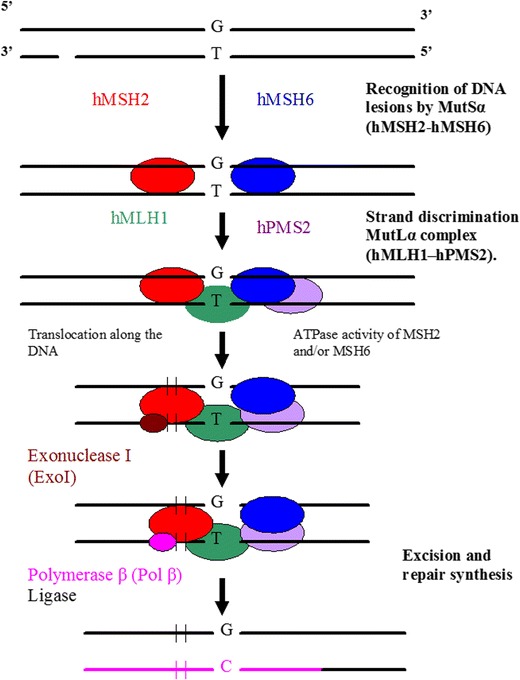
The mismatch repair (MMR). Recognition of DNA lesions occurs by heterodimer hMSH2–hMSH6 (MutSα). Upon binding to the mismatch, MutSα associates with heterodimeric complex hMLH1-hPMS2 (MutLα). After formation of a complex composed of MutSα and MutLα, excision is performed by ExoI and repair synthesis by Polβ

The homologous recombination (HR) system. HR starts with nucleolytic resection of the double strand break in the 5′ → 3′ direction by the MRE11–Rad50–NBS1 (MRN) complex, forming a 3′ single-stranded DNA fragment to which RAD52 homolog (S. cerevisiae) (Rad52) binds. Rad52 interacts with Rad51 homolog (RecA homolog, E. coli) (S. cerevisiae) (Rad51), provoking a DNA strand exchange with the undamaged, homologous DNA molecule. After DNA synthesis, ligation and branch migration, the resulting structure is resolved

The non-homologous end joining (NHEJ). The first step in NHEJ is the binding of a heterodimeric complex consisting of the proteins Ku70 and Ku80. Ku heterodimer associates with the catalytic subunit of DNA-dependent protein kinase (DNA–PKcs), forming the DNA–PK holoenzyme. One of the targets of DNA–PKcs is X-ray repair cross-complementing protein 4 (XRCC4). XRCC4 forms a stable complex with DNA ligase IV. DNA–PK activates XRCC4–ligase IV, which links the broken DNA ends together. Before re-ligation by XRCC4–ligase IV, the DNA ends are processed by the MRE11–Rad50–NBS1 complex, presumably involving Artemis
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
