

The role of juvenile hormone in dominance behavior, reproduction and cuticular pheromone signaling in the caste-flexible epiponine wasp, Synoeca surinama
- PMID: 25371699
- PMCID: PMC4219083
- DOI: 10.1186/s12983-014-0078-5
The role of juvenile hormone in dominance behavior, reproduction and cuticular pheromone signaling in the caste-flexible epiponine wasp, Synoeca surinama
Abstract
Background: The popular view on insect sociality is that of a harmonious division of labor among two morphologically distinct and functionally non-overlapping castes. But this is a highly derived state and not a prerequisite for a functional society. Rather, caste-flexibility is a central feature in many eusocial wasps, where adult females have the potential to become queens or workers, depending on the social environment. In non-swarming paper wasps (e.g., Polistes), prospective queens fight one another to assert their dominance, with losers becoming workers if they remain on the nest. This aggression is fueled by juvenile hormone (JH) and ecdysteroids, major factors involved in caste differentiation in most eusocial insects. We tested whether these hormones have conserved aggression-promoting functions in Synoeca surinama, a caste-flexible swarm-founding wasp (Epiponini) where reproductive competition is high and aggressive displays are common.
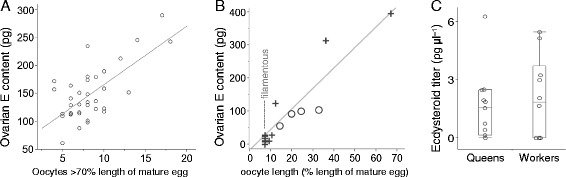
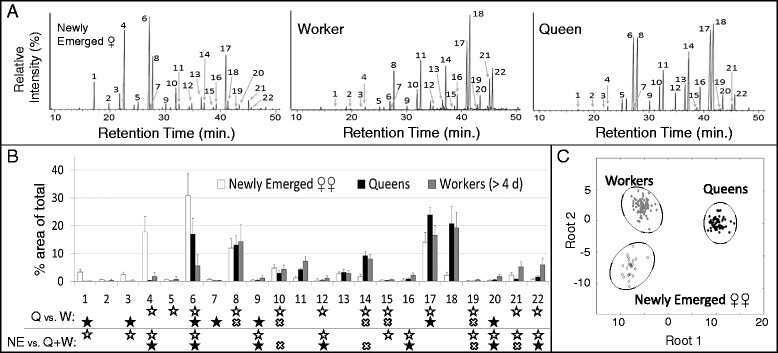
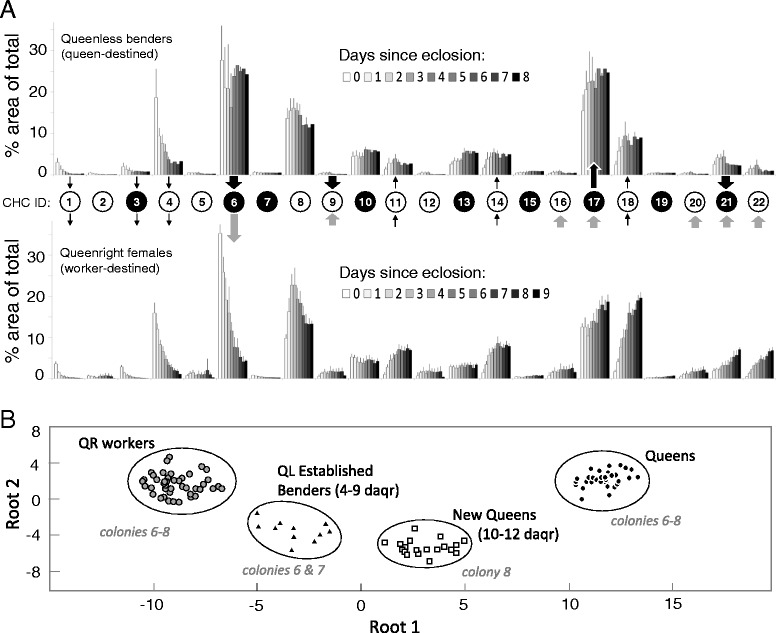
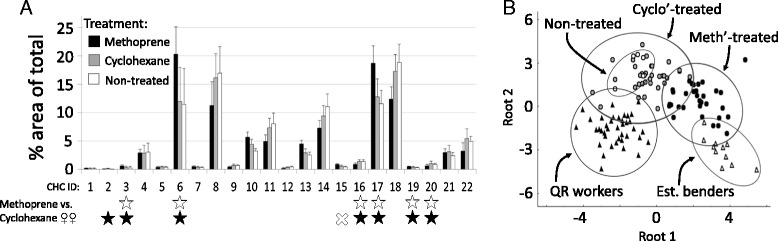
Results: We observed the behavioral interactions of S. surinama females in field nests before and after we had removed the egg-laying queen(s). We measured the ovarian reproductive status, hemolymph JH and ecdysteroid titers, ovarian ecdysteroid content, and analyzed the cuticular hydrocarbon (CHC) composition of females engaged in competitive interactions in both queenright and queenless contexts. These data, in combination with hormone manipulation experiments, revealed that neither JH nor ecdysteroids are necessary for the expression of dominance behaviors in S. surinama. Instead, we show that JH likely functions as a gonadotropin and directly modifies the cuticular hydrocarbon blend of young workers to match that of a reproductive. Hemolymph ecdysteroids, in contrast, are not different between queens and workers despite great differences in ovarian ecdysteroid content.
Conclusions: The endocrine profile of S. surinama shows surprising differences from those of other caste-flexible wasps, although a rise in JH titers in replacement queens is a common theme. Extensive remodeling of hormone functions is also evident in the highly eusocial bees, which has been attributed to the evolution of morphologically defined castes. Our results show that hormones which regulate caste-plasticity can lose these roles even while caste-plasticity is preserved.
Keywords: Cuticular hydrocarbons; Ecdysteroids; Endocrine; Epiponini; Juvenile hormone; Swarm founding; Wasps.
Figures




References
-
- West-Eberhard MJ. Intragroup selection and the evolution of insect societies. In: Alexander RD, Tinkle DW, editors. Natural Selection and Social Behavior. New York: Chiron Press; 1981. pp. 3–17.
-
- West-Eberhard MJ. The Establishment of Dominance of the Queen in Social Wasp Colonies. Netherlands: Wageningen; 1977. pp. 223–227.
-
- Platt TG, Queller DC, Strassmann JE. Aggression and worker control of caste fate in a multiple-queen wasp, Parachartergus colobopterus. Anim Behav. 2004;67:1–10. doi: 10.1016/j.anbehav.2003.01.005. - DOI
-
- West-Eberhard MJ. The nature and evolution of swarming in tropical social wasps (Vespidae, Polistinae, Polybiini) In: Jaisson P, editor. Social Insects in the Tropics. Paris: University of Paris Press; 1982. pp. 97–128.
LinkOut - more resources
Full Text Sources
Other Literature Sources
