

ESCRT-II/Vps25 constrains digit number by endosome-mediated selective modulation of FGF-SHH signaling
- PMID: 25373905
- PMCID: PMC4223648
- DOI: 10.1016/j.celrep.2014.09.019
ESCRT-II/Vps25 constrains digit number by endosome-mediated selective modulation of FGF-SHH signaling
Abstract
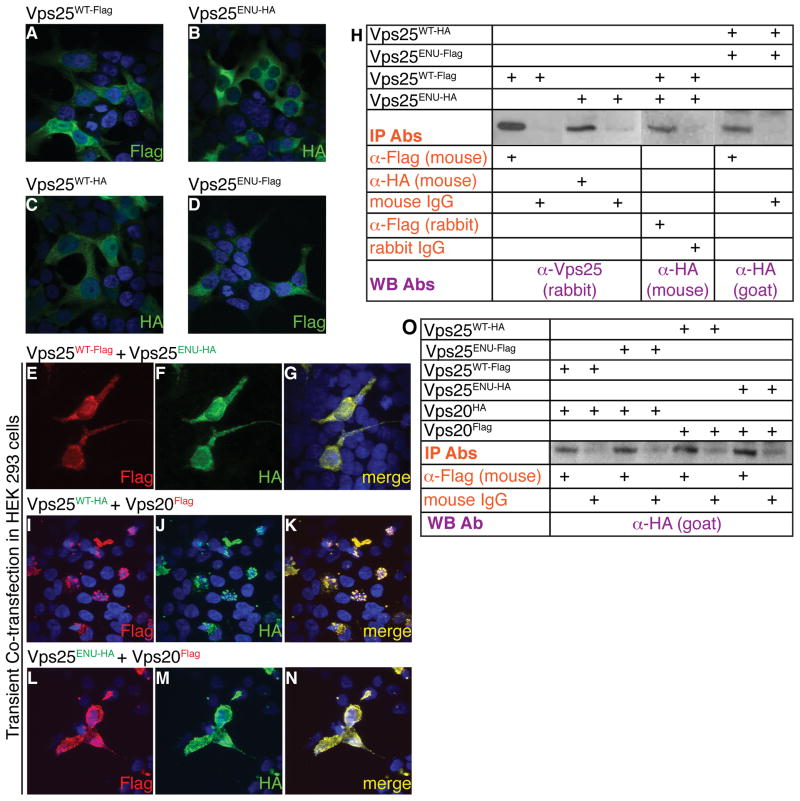
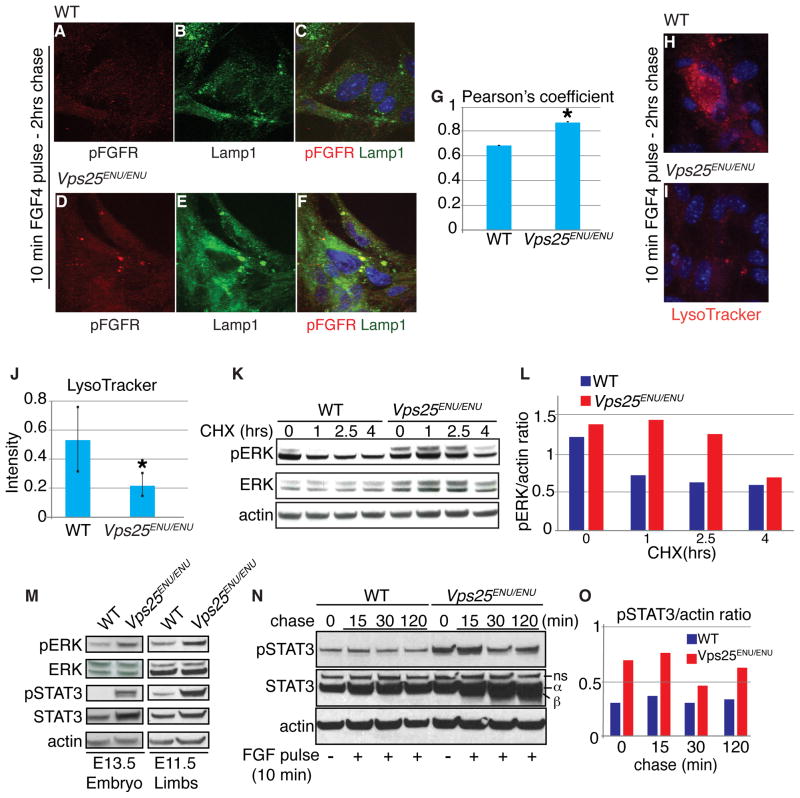
Sorting and degradation of receptors and associated signaling molecules maintain homeostasis of conserved signaling pathways during cell specification and tissue development. Yet, whether machineries that sort signaling proteins act preferentially on different receptors and ligands in different contexts remains mysterious. Here, we show that Vacuolar protein sorting 25, Vps25, a component of ESCRT-II (Endosomal Sorting Complex Required for Transport II), directs preferential endosome-mediated modulation of FGF signaling in limbs. By ENU-induced mutagenesis, we isolated a polydactylous mouse line carrying a hypomorphic mutation of Vps25 (Vps25(ENU)). Unlike Vps25-null embryos we generated, Vps25(ENU/ENU) mutants survive until late gestation. Their limbs display FGF signaling enhancement and consequent hyperactivation of the FGF-SHH feedback loop causing polydactyly, whereas WNT and BMP signaling remain unperturbed. Notably, Vps25(ENU/ENU) Mouse Embryonic Fibroblasts exhibit aberrant FGFR trafficking and degradation; however, SHH signaling is unperturbed. These studies establish that the ESCRT-II machinery selectively limits FGF signaling in vertebrate skeletal patterning.
Figures





References
-
- Anderson E, Peluso S, Lettice LA, Hill RE. Human limb abnormalities caused by disruption of hedgehog signaling. Trends Genet. 2012;28:364–373. - PubMed
-
- Anderson KV, Ingham PW. The transformation of the model organism: a decade of developmental genetics. Nat Genet. 2003;33:285–293. - PubMed
-
- Babst M, Katzmann DJ, Estepa-Sabal EJ, Meerloo T, Emr SD. Escrt-III: an endosome-associated heterooligomeric protein complex required for mvb sorting. Dev Cell. 2002a;3:271–282. - PubMed
-
- Babst M, Katzmann DJ, Snyder WB, Wendland B, Emr SD. Endosome-associated complex, ESCRT-II, recruits transport machinery for protein sorting at the multivesicular body. Dev Cell. 2002b;3:283–289. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- U42 RR024244/RR/NCRR NIH HHS/United States
- 2R01HD043997/HD/NICHD NIH HHS/United States
- R21DE018031/DE/NIDCR NIH HHS/United States
- U01HG004085/HG/NHGRI NIH HHS/United States
- R21DE018031-02S1/DE/NIDCR NIH HHS/United States
- R01 HD043997/HD/NICHD NIH HHS/United States
- P30 CA008748/CA/NCI NIH HHS/United States
- R21 DE018031/DE/NIDCR NIH HHS/United States
- R37 HD035455/HD/NICHD NIH HHS/United States
- U01 HG004080/HG/NHGRI NIH HHS/United States
- U42RR024244/RR/NCRR NIH HHS/United States
- U01HG004080/HG/NHGRI NIH HHS/United States
- T32 GM007739/GM/NIGMS NIH HHS/United States
- U01 HG004085/HG/NHGRI NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
