

Pupil fluctuations track fast switching of cortical states during quiet wakefulness
- PMID: 25374359
- PMCID: PMC4323337
- DOI: 10.1016/j.neuron.2014.09.033
Pupil fluctuations track fast switching of cortical states during quiet wakefulness
Abstract
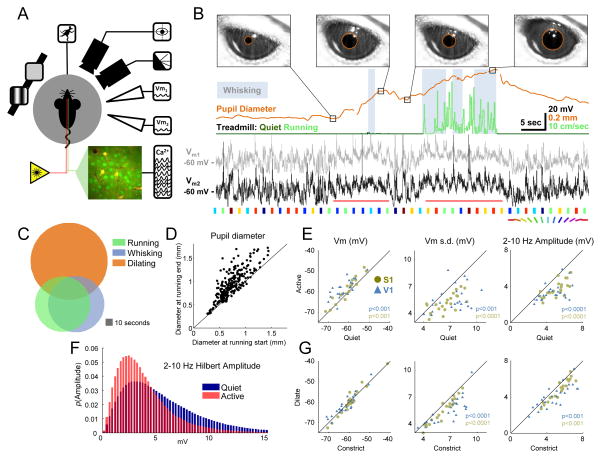
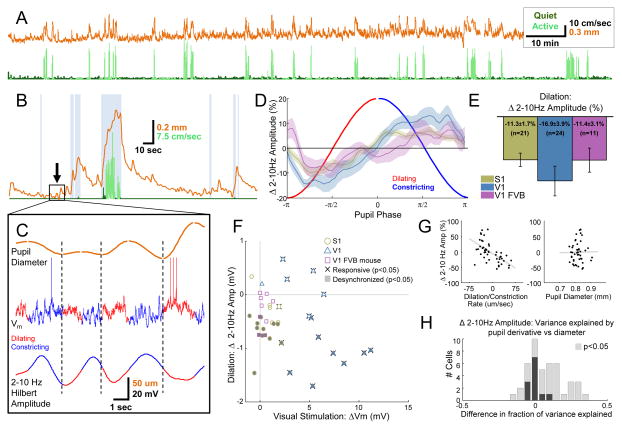
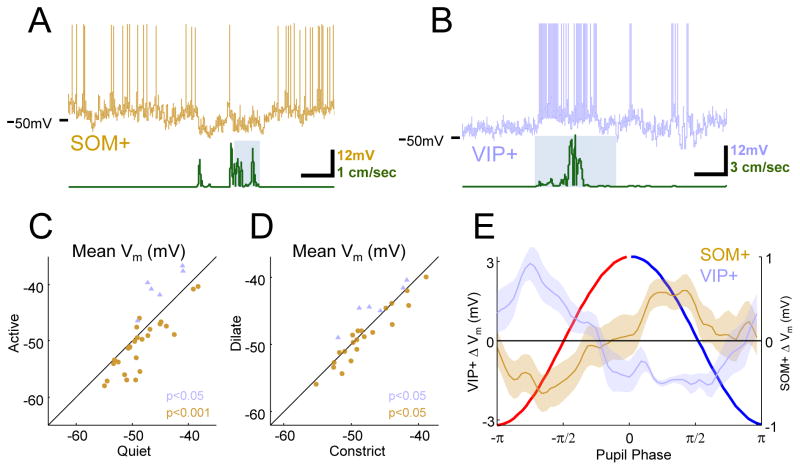
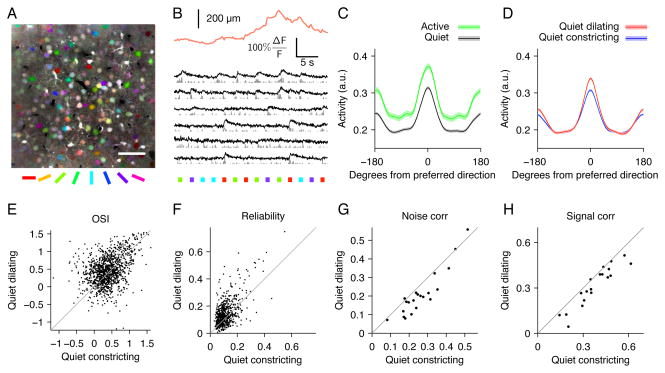
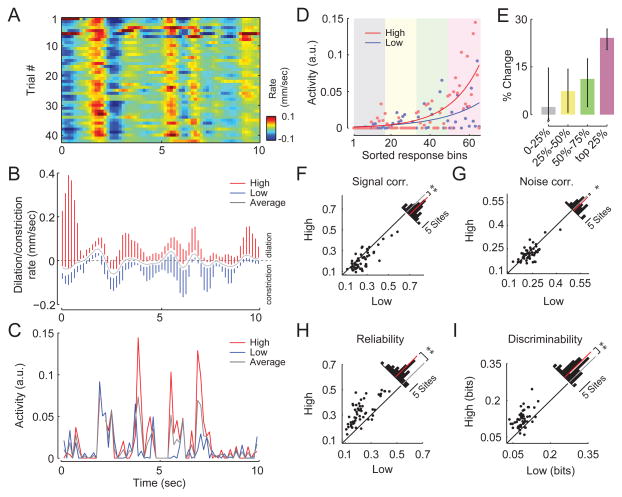
Neural responses are modulated by brain state, which varies with arousal, attention, and behavior. In mice, running and whisking desynchronize the cortex and enhance sensory responses, but the quiescent periods between bouts of exploratory behaviors have not been well studied. We found that these periods of "quiet wakefulness" were characterized by state fluctuations on a timescale of 1-2 s. Small fluctuations in pupil diameter tracked these state transitions in multiple cortical areas. During dilation, the intracellular membrane potential was desynchronized, sensory responses were enhanced, and population activity was less correlated. In contrast, constriction was characterized by increased low-frequency oscillations and higher ensemble correlations. Specific subtypes of cortical interneurons were differentially activated during dilation and constriction, consistent with their participation in the observed state changes. Pupillometry has been used to index attention and mental effort in humans, but the intracellular dynamics and differences in population activity underlying this phenomenon were previously unknown.
Figures
Comment in
-
Keeping an eye on cortical states.Neuron. 2014 Oct 22;84(2):246-8. doi: 10.1016/j.neuron.2014.10.005. Epub 2014 Oct 22. Neuron. 2014. PMID: 25374350
References
-
- Aston-Jones G, Rajkowski J, Cohen J. Role of locus coeruleus in attention and behavioral flexibility. Biological Psychiatry. 1999;46:1309–1320. - PubMed
-
- Chang B, Hawes NL, Hurd RE, Davisson MT, Nusinowitz S, Heckenlively JR. Retinal degeneration mutants in the mouse. Vision Res. 2002;42:517–525. - PubMed
Publication types
MeSH terms
Grants and funding
- T32EY07001/EY/NEI NIH HHS/United States
- F30 MH095440/MH/NIMH NIH HHS/United States
- F30MH095440/MH/NIMH NIH HHS/United States
- U54 HD083092/HD/NICHD NIH HHS/United States
- DP1 OD008301/OD/NIH HHS/United States
- DP1EY023176/DP/NCCDPHP CDC HHS/United States
- T32 EY007001/EY/NEI NIH HHS/United States
- T32GM007330/GM/NIGMS NIH HHS/United States
- T32EB006350/EB/NIBIB NIH HHS/United States
- P30 EY002520/EY/NEI NIH HHS/United States
- T32 EB006350/EB/NIBIB NIH HHS/United States
- P30EY002520/EY/NEI NIH HHS/United States
- T32 GM007330/GM/NIGMS NIH HHS/United States
- DP1 EY023176/EY/NEI NIH HHS/United States
- DP1OD008301/OD/NIH HHS/United States
- R01 DA028525/DA/NIDA NIH HHS/United States
- R01DA028525/DA/NIDA NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
