

Engagement of neural circuits underlying 2D spatial navigation in a rodent virtual reality system
- PMID: 25374363
- PMCID: PMC4454359
- DOI: 10.1016/j.neuron.2014.08.042
Engagement of neural circuits underlying 2D spatial navigation in a rodent virtual reality system
Abstract
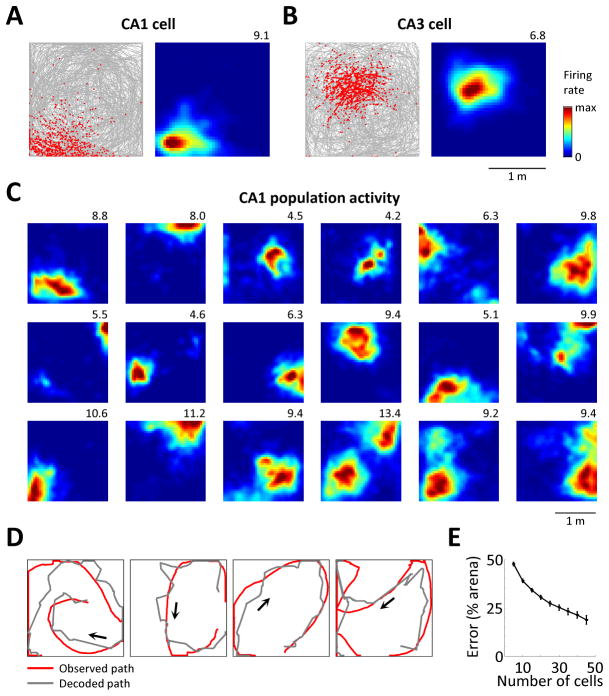
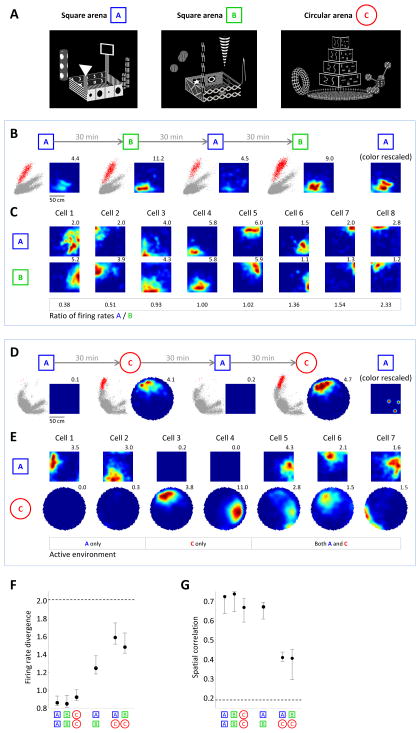
Virtual reality (VR) enables precise control of an animal's environment and otherwise impossible experimental manipulations. Neural activity in rodents has been studied on virtual 1D tracks. However, 2D navigation imposes additional requirements, such as the processing of head direction and environment boundaries, and it is unknown whether the neural circuits underlying 2D representations can be sufficiently engaged in VR. We implemented a VR setup for rats, including software and large-scale electrophysiology, that supports 2D navigation by allowing rotation and walking in any direction. The entorhinal-hippocampal circuit, including place, head direction, and grid cells, showed 2D activity patterns similar to those in the real world. Furthermore, border cells were observed, and hippocampal remapping was driven by environment shape, suggesting functional processing of virtual boundaries. These results illustrate that 2D spatial representations can be engaged by visual and rotational vestibular stimuli alone and suggest a novel VR tool for studying rat navigation.
Figures





References
-
- Aflalo TN, Graziano MS. Four-dimensional spatial reasoning in humans. Journal of experimental psychology Human perception and performance. 2008;34:1066–1077. - PubMed
-
- Barry C, Hayman R, Burgess N, Jeffery KJ. Experience-dependent rescaling of entorhinal grids. Nature neuroscience. 2007;10:682–684. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
